

RLIP76 (RalBP1) is an R-Ras effector that mediates adhesion-dependent Rac activation and cell migration
- PMID: 16966426
- PMCID: PMC2064341
- DOI: 10.1083/jcb.200603111
RLIP76 (RalBP1) is an R-Ras effector that mediates adhesion-dependent Rac activation and cell migration
Abstract
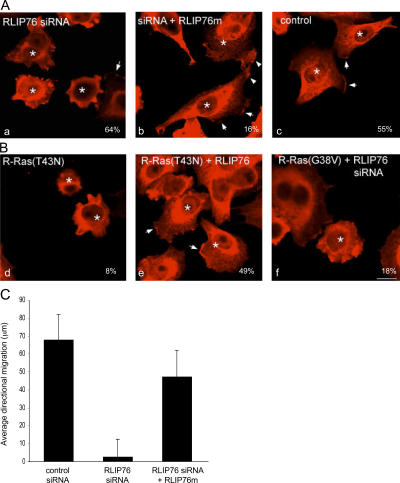
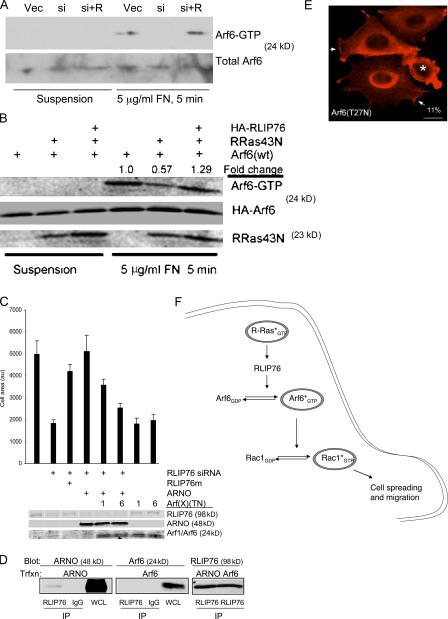
The Ras family of small GTPases regulates cell proliferation, spreading, migration and apoptosis, and malignant transformation by binding to several protein effectors. One such GTPase, R-Ras, plays distinct roles in each of these processes, but to date, identified R-Ras effectors were shared with other Ras family members (e.g., H-Ras). We utilized a new database of Ras-interacting proteins to identify RLIP76 (RalBP1) as a novel R-Ras effector. RLIP76 binds directly to R-Ras in a GTP-dependent manner, but does not physically associate with the closely related paralogues H-Ras and Rap1A. RLIP76 is required for adhesion-induced Rac activation and the resulting cell spreading and migration, as well as for the ability of R-Ras to enhance these functions. RLIP76 regulates Rac activity through the adhesion-induced activation of Arf6 GTPase and activation of Arf6 bypasses the requirement for RLIP76 in Rac activation and cell spreading. Thus, we identify a novel R-Ras effector, RLIP76, which links R-Ras to adhesion-induced Rac activation through a GTPase cascade that mediates cell spreading and migration.
Figures
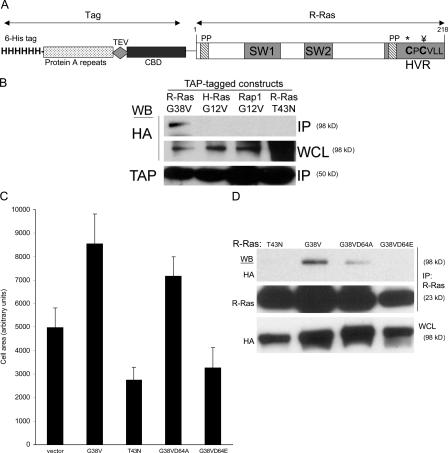
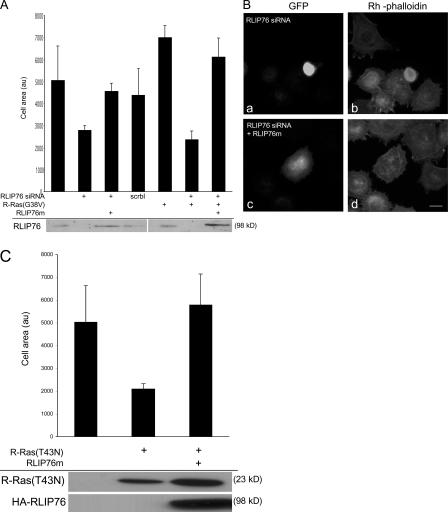
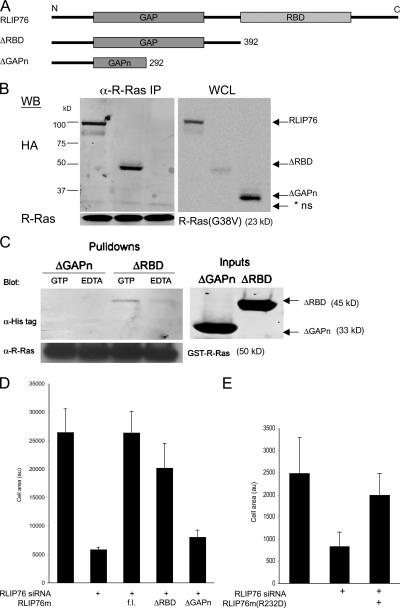
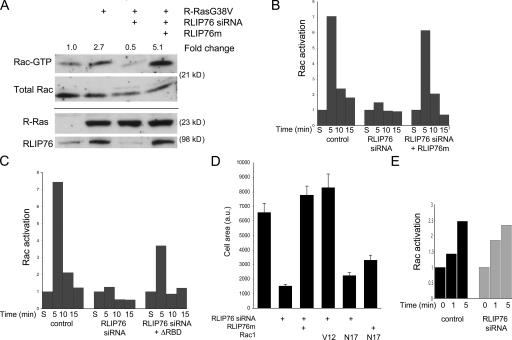
References
-
- Ada-Nguema, A.S., H. Xenias, M.P. Sheetz, and P.J. Keely. 2006. The small GTPase R-Ras regulates organization of actin and drives membrane protrusions through the activity of PLC{epsilon}. J. Cell Sci. 119:1307–1319. - PubMed
-
- Bagrodia, S., S.J. Taylor, K.A. Jordon, A.L. Van, and R.A. Cerione. 1998. A novel regulator of p21-activated kinases. J. Biol. Chem. 273:23633–23636. - PubMed
-
- Bauer, B., G. Mirey, I.R. Vetter, J.A. Garcia-Ranea, A. Valencia, A. Wittinghofer, J.H. Camonis, and R.H. Cool. 1999. Effector recognition by the small GTP-binding proteins Ras and Ral. J. Biol. Chem. 274:17763–17770. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
