

Continuous recruitment of naive T cells contributes to heterogeneity of antiviral CD8 T cells during persistent infection
- PMID: 16966427
- PMCID: PMC2118117
- DOI: 10.1084/jem.20060995
Continuous recruitment of naive T cells contributes to heterogeneity of antiviral CD8 T cells during persistent infection
Abstract
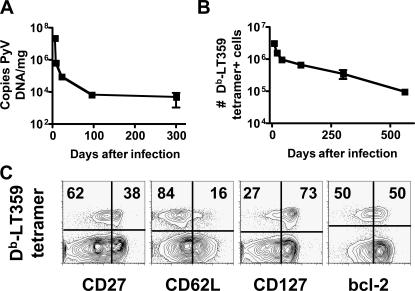
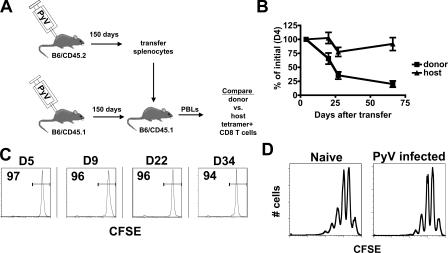
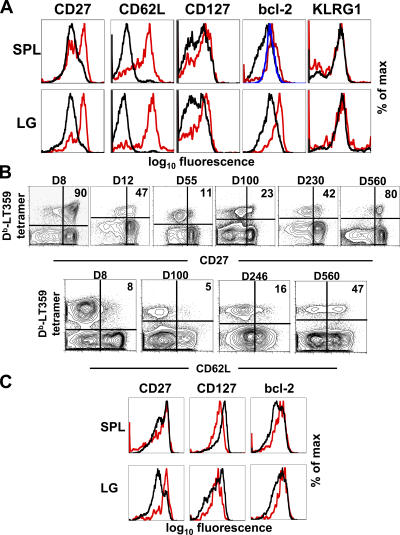
Numerous microbes establish persistent infections, accompanied by antigen-specific CD8 T cell activation. Pathogen-specific T cells in chronically infected hosts are often phenotypically and functionally variable, as well as distinct from T cells responding to nonpersistent infections; this phenotypic heterogeneity has been attributed to an ongoing reencounter with antigen. Paradoxically, maintenance of memory CD8 T cells to acutely resolved infections is antigen independent, whereas there is a dependence on antigen for T cell survival in chronically infected hosts. Using two chronic viral infections, we demonstrate that new naive antigen-specific CD8 T cells are primed after the acute phase of infection. These newly recruited T cells are phenotypically distinct from those primed earlier. Long-lived antiviral CD8 T cells are defective in self-renewal, and lack of thymic output results in the decline of virus-specific CD8 T cells, indicating that newly generated T cells preserve antiviral CD8 T cell populations during chronic infection. These findings reveal a novel role for antigen in maintaining virus-specific CD8 T cells during persistent infection and provide insight toward understanding T cell differentiation in chronic infection.
Figures

Similar articles
-
Viral antigen and extensive division maintain virus-specific CD8 T cells during chronic infection.J Exp Med. 2007 Apr 16;204(4):941-9. doi: 10.1084/jem.20061937. Epub 2007 Apr 9. J Exp Med. 2007. PMID: 17420267 Free PMC article.
-
Differential role of IL-2R signaling for CD8+ T cell responses in acute and chronic viral infections.Eur J Immunol. 2007 Jun;37(6):1502-12. doi: 10.1002/eji.200637023. Eur J Immunol. 2007. PMID: 17492805
-
The role of CD80/CD86 in generation and maintenance of functional virus-specific CD8+ T cells in mice infected with lymphocytic choriomeningitis virus.J Immunol. 2010 Aug 1;185(3):1730-43. doi: 10.4049/jimmunol.0903894. Epub 2010 Jul 2. J Immunol. 2010. PMID: 20601595
-
Virus-specific CD8 T cells: activation, differentiation and memory formation.APMIS. 2009 May;117(5-6):356-81. doi: 10.1111/j.1600-0463.2009.02459.x. APMIS. 2009. PMID: 19400862 Review.
-
The architectural design of CD8+ T cell responses in acute and chronic infection: Parallel structures with divergent fates.J Exp Med. 2021 Apr 5;218(4):e20201730. doi: 10.1084/jem.20201730. J Exp Med. 2021. PMID: 33755719 Free PMC article. Review.
Cited by
-
T cells maintain an exhausted phenotype after antigen withdrawal and population reexpansion.Nat Immunol. 2013 Jun;14(6):603-10. doi: 10.1038/ni.2606. Epub 2013 May 5. Nat Immunol. 2013. PMID: 23644506
-
Allogeneic differences in the dependence on CD4+ T-cell help for virus-specific CD8+ T-cell differentiation.J Virol. 2007 Dec;81(24):13743-53. doi: 10.1128/JVI.01778-07. Epub 2007 Oct 3. J Virol. 2007. PMID: 17913814 Free PMC article.
-
Evidence of premature immune aging in patients thymectomized during early childhood.J Clin Invest. 2009 Oct;119(10):3070-8. doi: 10.1172/JCI39269. Epub 2009 Sep 21. J Clin Invest. 2009. PMID: 19770514 Free PMC article.
-
Autoimmune vitiligo does not require the ongoing priming of naive CD8 T cells for disease progression or associated protection against melanoma.J Immunol. 2014 Feb 15;192(4):1433-9. doi: 10.4049/jimmunol.1302139. Epub 2014 Jan 8. J Immunol. 2014. PMID: 24403535 Free PMC article.
-
De novo recruitment of antigen-experienced and naive T cells contributes to the long-term maintenance of antiviral T cell populations in the persistently infected central nervous system.J Immunol. 2009 Oct 15;183(8):5163-70. doi: 10.4049/jimmunol.0902164. Epub 2009 Sep 28. J Immunol. 2009. PMID: 19786545 Free PMC article.
References
-
- Schmitz, J.E., M.J. Kuroda, S. Santra, V.G. Sasseville, M.A. Simon, M.A. Lifton, P. Racz, K. Tenner-Racz, M. Dalesandro, B.J. Scallon, et al. 1999. Control of viremia in simian immunodeficiency virus infection by CD8+ lymphocytes. Science. 283:857–860. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
