

Cellular effects and metabolic stability of N1-cyclic inosine diphosphoribose and its derivatives
- PMID: 16967053
- PMCID: PMC1978434
- DOI: 10.1038/sj.bjp.0706869
Cellular effects and metabolic stability of N1-cyclic inosine diphosphoribose and its derivatives
Abstract
Background and purpose: Recently, a number of mimics of the second messenger cyclic ADP-ribose (cADPR) with replacement of adenosine by inosine were introduced. In addition, various alterations in the molecule ranging from substitutions at C8 of the base up to full replacement of the ribose moieties still retained biological activity. However, nothing is known about the metabolic stability and cellular effects of these novel analogues.
Experimental approach: cADPR and the inosine-based analogues were incubated with CD38, ADP-ribosyl cyclase and NAD-glycohydrolase and metabolism was analysed by RP-HPLC. Furthermore, the effect of the analogues on cytokine expression and proliferation was investigated in primary T-lymphocytes and T-lymphoma cells.
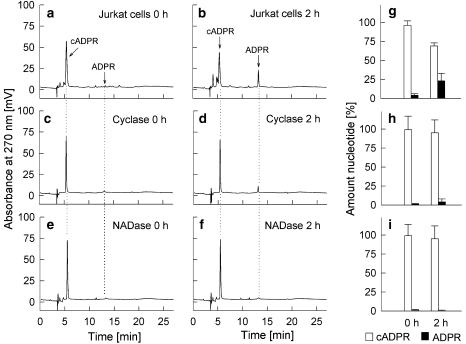
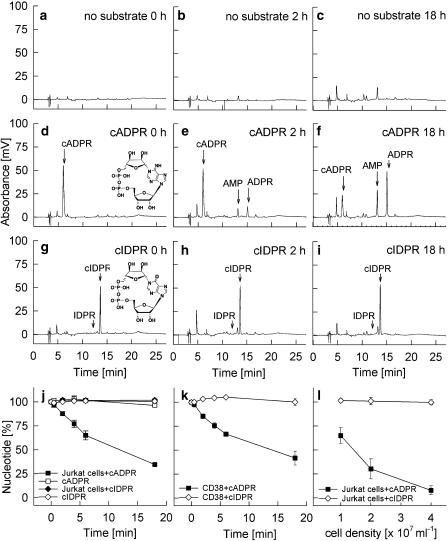
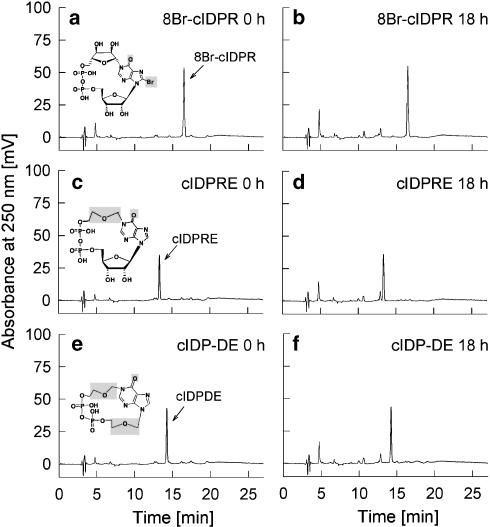
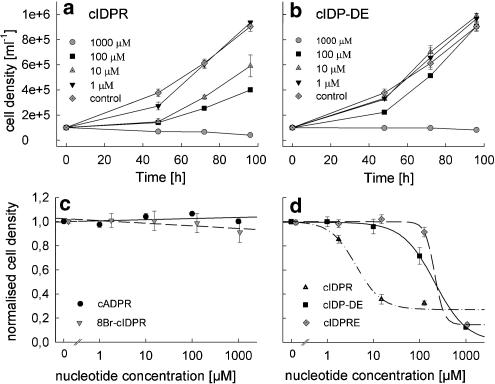
Key results: Incubation of cADPR with CD38 resulted in degradation to adenosine diphosphoribose. ADP-ribosyl cyclase weakly catabolised cADPR whereas NAD-glycohydrolase showed no such activity. In contrast, N1-cyclic inosine 5'-diphosphoribose (N1-cIDPR) was not hydrolyzed by CD38. Three additional N1-cIDPR analogues showed a similar stability. Proliferation of Jurkat T-lymphoma cells was inhibited by N1-cIDPR, N1-[(phosphoryl-O-ethoxy)-methyl]-N9-[(phosphoryl-O-ethoxy)-methyl]-hypoxanthine-cyclic pyrophosphate (N1-cIDP-DE) and N1-ethoxymethyl-cIDPR (N1-cIDPRE). In contrast, in primary T cells neither proliferation nor cytokine expression was affected by these compounds.
Conclusions and implications: The metabolic stability of N1-cIDPR and its analogues provides an advantage for the development of novel pharmaceutical compounds interfering with cADPR mediated Ca2+ signalling pathways. The differential effects of N1-cIDPR and N1-cIDPRE on proliferation and cytokine expression in primary T cells versus T-lymphoma cells may constitute a starting point for novel anti-tumor drugs.
Figures
Similar articles
-
8-Bromo-cyclic inosine diphosphoribose: towards a selective cyclic ADP-ribose agonist.Biochem J. 2009 Jul 29;422(1):139-49. doi: 10.1042/BJ20082308. Biochem J. 2009. PMID: 19492987 Free PMC article.
-
Aberrant cyclization affords a C-6 modified cyclic adenosine 5'-diphosphoribose analogue with biological activity in Jurkat T cells.J Med Chem. 2012 Feb 23;55(4):1478-89. doi: 10.1021/jm201127y. Epub 2012 Feb 8. J Med Chem. 2012. PMID: 22248391 Free PMC article.
-
Structural determinants for N1/N7 cyclization of nicotinamide hypoxanthine 5'-dinucleotide (NHD+) derivatives by ADP-ribosyl cyclase from aplysia californica: Ca2+-mobilizing activity of 8-substituted cyclic inosine 5'-diphosphoribose analogues in T-lymphocytes.J Med Chem. 2006 Aug 24;49(17):5162-76. doi: 10.1021/jm060275a. J Med Chem. 2006. PMID: 16913705
-
Biochemistry, biology, and pharmacology of cyclic adenosine diphosphoribose (cADPR).Curr Med Chem. 2004 Apr;11(7):847-55. doi: 10.2174/0929867043455602. Curr Med Chem. 2004. PMID: 15078169 Review.
-
Autocrine and paracrine calcium signaling by the CD38/NAD+/cyclic ADP-ribose system.Ann N Y Acad Sci. 2004 Dec;1028:176-91. doi: 10.1196/annals.1322.021. Ann N Y Acad Sci. 2004. PMID: 15650244 Review.
Cited by
-
cADPR Does Not Activate TRPM2.Int J Mol Sci. 2022 Mar 15;23(6):3163. doi: 10.3390/ijms23063163. Int J Mol Sci. 2022. PMID: 35328585 Free PMC article.
-
A novel fluorescent cell membrane-permeable caged cyclic ADP-ribose analogue.J Biol Chem. 2012 Jul 13;287(29):24774-83. doi: 10.1074/jbc.M111.329854. Epub 2012 Jun 1. J Biol Chem. 2012. PMID: 22661714 Free PMC article.
-
CD38 Structure-Based Inhibitor Design Using the N1-Cyclic Inosine 5'-Diphosphate Ribose Template.PLoS One. 2013 Jun 19;8(6):e66247. doi: 10.1371/journal.pone.0066247. Print 2013. PLoS One. 2013. PMID: 23840430 Free PMC article.
-
8-Bromo-cyclic inosine diphosphoribose: towards a selective cyclic ADP-ribose agonist.Biochem J. 2009 Jul 29;422(1):139-49. doi: 10.1042/BJ20082308. Biochem J. 2009. PMID: 19492987 Free PMC article.
-
Second messenger analogues highlight unexpected substrate sensitivity of CD38: total synthesis of the hybrid "L-cyclic inosine 5'-diphosphate ribose".Sci Rep. 2017 Nov 23;7(1):16100. doi: 10.1038/s41598-017-16388-0. Sci Rep. 2017. PMID: 29170518 Free PMC article.
References
-
- Bailey VC, Fortt SM, Summerhill RJ, Galione A, Potter BVL. Cyclic aristeromycin diphosphate ribose: a potent and poorly hydrolysable Ca(2+)-mobilising mimic of cyclic adenosine diphosphate ribose. FEBS Lett. 1996;379:227–230. - PubMed
-
- Bruzzone S, Guida L, Zocchi E, Franco L, De Flora A. Connexin 43 hemi channels mediate Ca2+-regulated transmembrane NAD+ fluxes in intact cells. FASEB J. 2001;15:10–12. - PubMed
-
- Da Silva CP, Schweitzer K, Heyer P, Malavasi F, Mayr GW, Guse AH. Ectocellular CD38-catalyzed synthesis and intracellular Ca2+-signalling activity of cyclic ADP-ribose in T-lymphocytes are not functionally related. FEBS Lett. 1998;439:291–296. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
