

Threshold-dependent BMP-mediated repression: a model for a conserved mechanism that patterns the neuroectoderm
- PMID: 16968133
- PMCID: PMC1563485
- DOI: 10.1371/journal.pbio.0040313
Threshold-dependent BMP-mediated repression: a model for a conserved mechanism that patterns the neuroectoderm
Abstract
Subdivision of the neuroectoderm into three rows of cells along the dorsal-ventral axis by neural identity genes is a highly conserved developmental process. While neural identity genes are expressed in remarkably similar patterns in vertebrates and invertebrates, previous work suggests that these patterns may be regulated by distinct upstream genetic pathways. Here we ask whether a potential conserved source of positional information provided by the BMP signaling contributes to patterning the neuroectoderm. We have addressed this question in two ways: First, we asked whether BMPs can act as bona fide morphogens to pattern the Drosophila neuroectoderm in a dose-dependent fashion, and second, we examined whether BMPs might act in a similar fashion in patterning the vertebrate neuroectoderm. In this study, we show that graded BMP signaling participates in organizing the neural axis in Drosophila by repressing expression of neural identity genes in a threshold-dependent fashion. We also provide evidence for a similar organizing activity of BMP signaling in chick neural plate explants, which may operate by the same double negative mechanism that acts earlier during neural induction. We propose that BMPs played an ancestral role in patterning the metazoan neuroectoderm by threshold-dependent repression of neural identity genes.
Conflict of interest statement
Competing interests. The authors have declared that no competing interests exist.
Figures
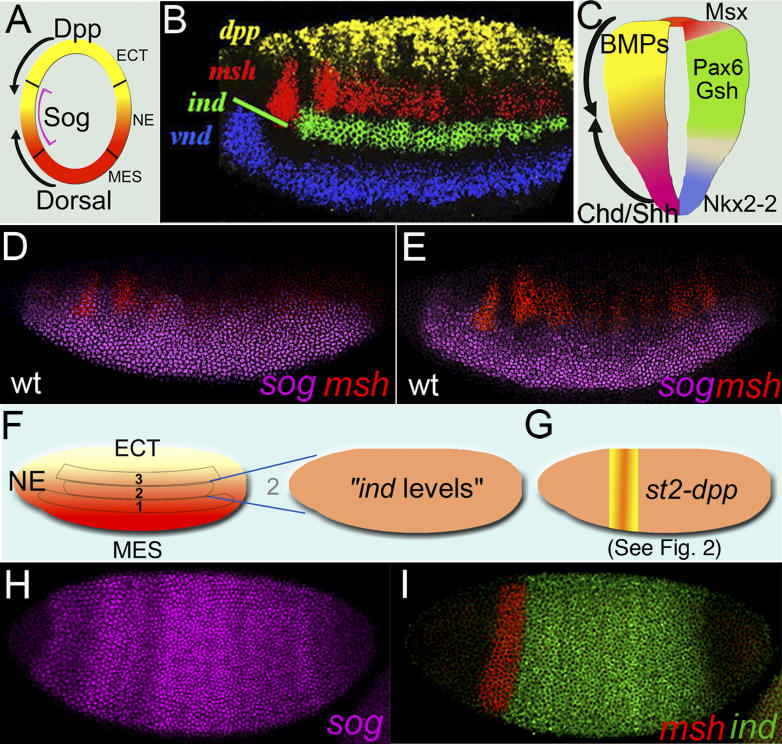
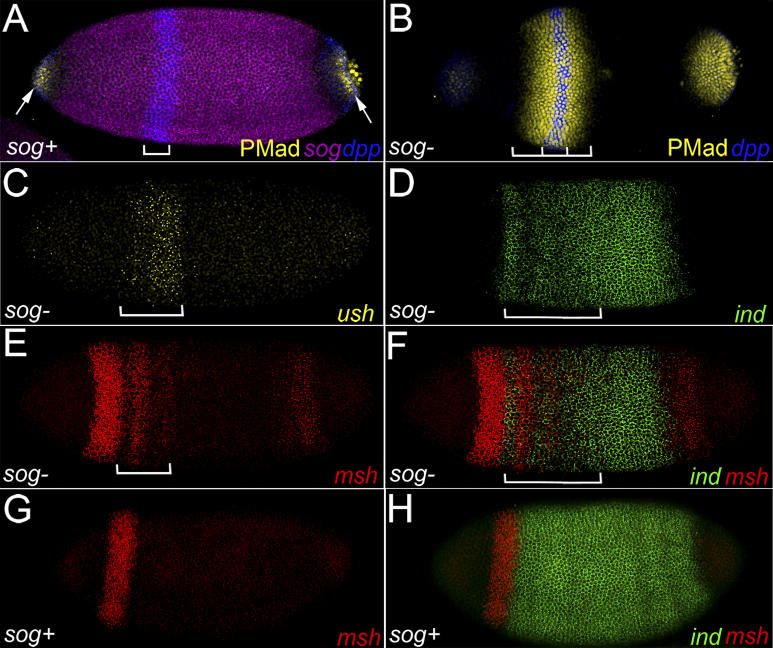
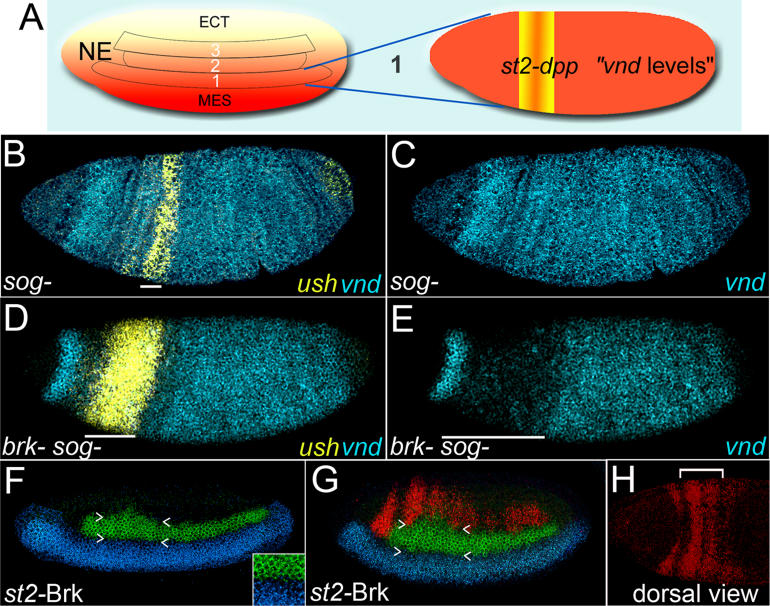
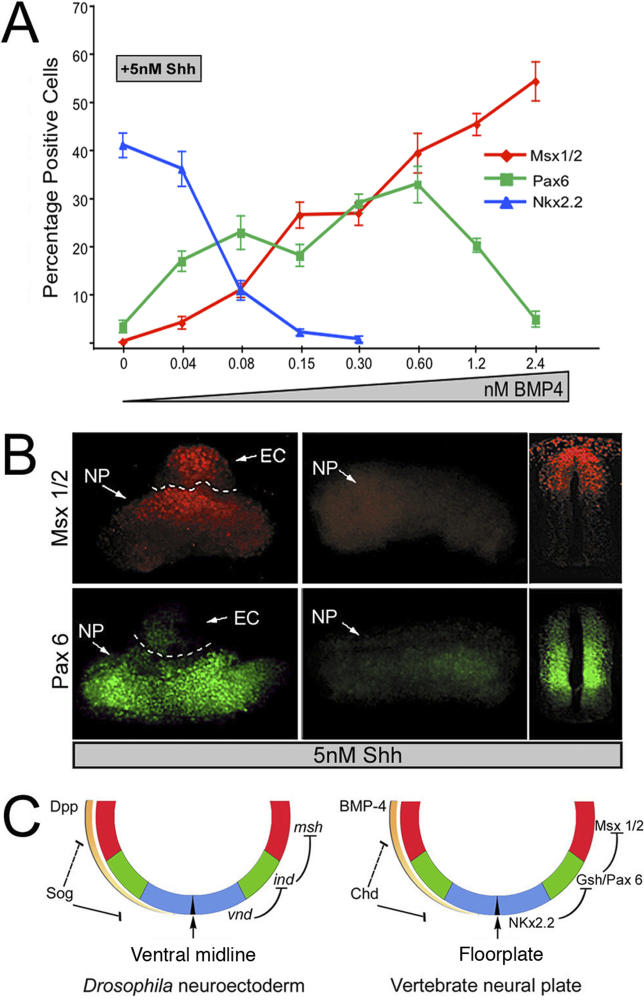
Comment in
-
BMPs: conserved morphogens in neural patterning.PLoS Biol. 2006 Oct;4(10):e346. doi: 10.1371/journal.pbio.0040346. Epub 2006 Sep 12. PLoS Biol. 2006. PMID: 20076475 Free PMC article. No abstract available.
References
-
- Freeman M, Gurdon JB. Regulatory principles of developmental signaling. Annu Rev Cell Dev Biol. 2002;18:515–539. - PubMed
-
- Tabata T, Takei Y. Morphogens, their identification and regulation. Development. 2004;131:703–712. - PubMed
-
- Bier E. Anti-neural-inhibition: A conserved mechanism for neural induction. Cell. 1997;89:681–684. - PubMed
-
- Eldar A, Dorfman R, Weiss D, Ashe H, Shilo BZ, et al. Robustness of the BMP morphogen gradient in Drosophila embryonic patterning. Nature. 2002;419:304–308. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
