

Diseases caused by defects in the visual cycle: retinoids as potential therapeutic agents
- PMID: 16968212
- PMCID: PMC2442882
- DOI: 10.1146/annurev.pharmtox.47.120505.105225
Diseases caused by defects in the visual cycle: retinoids as potential therapeutic agents
Abstract
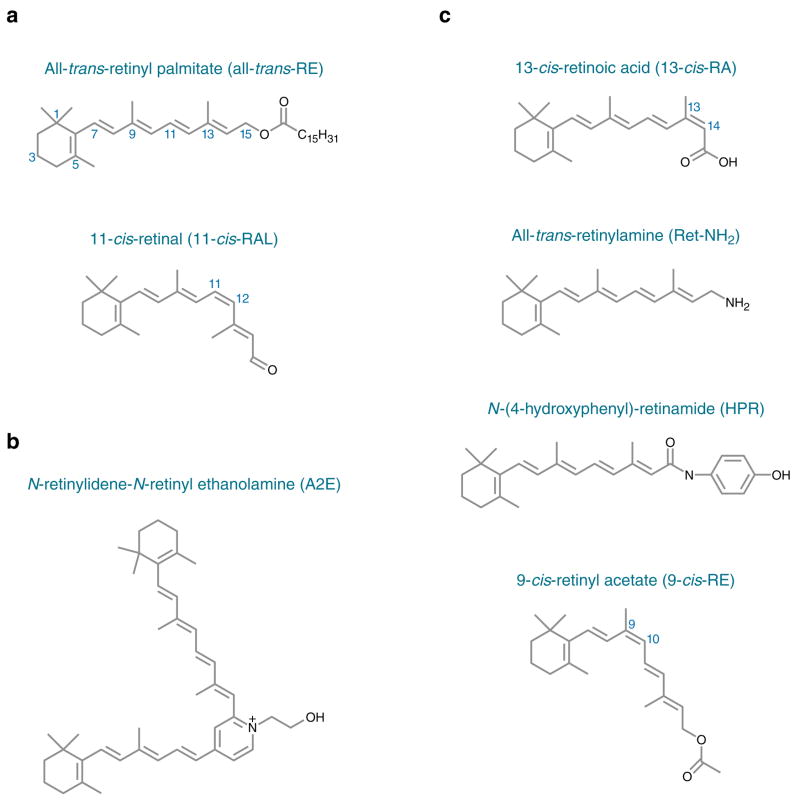
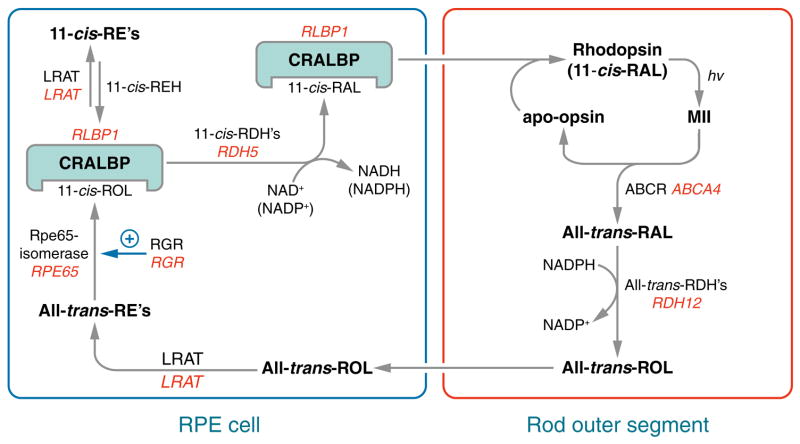
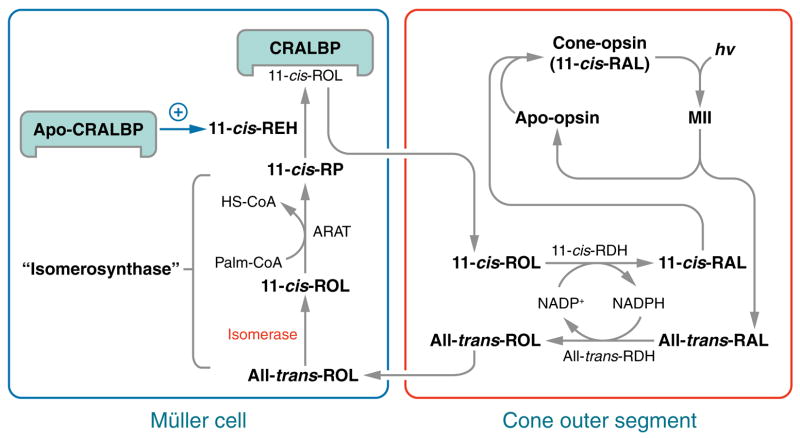
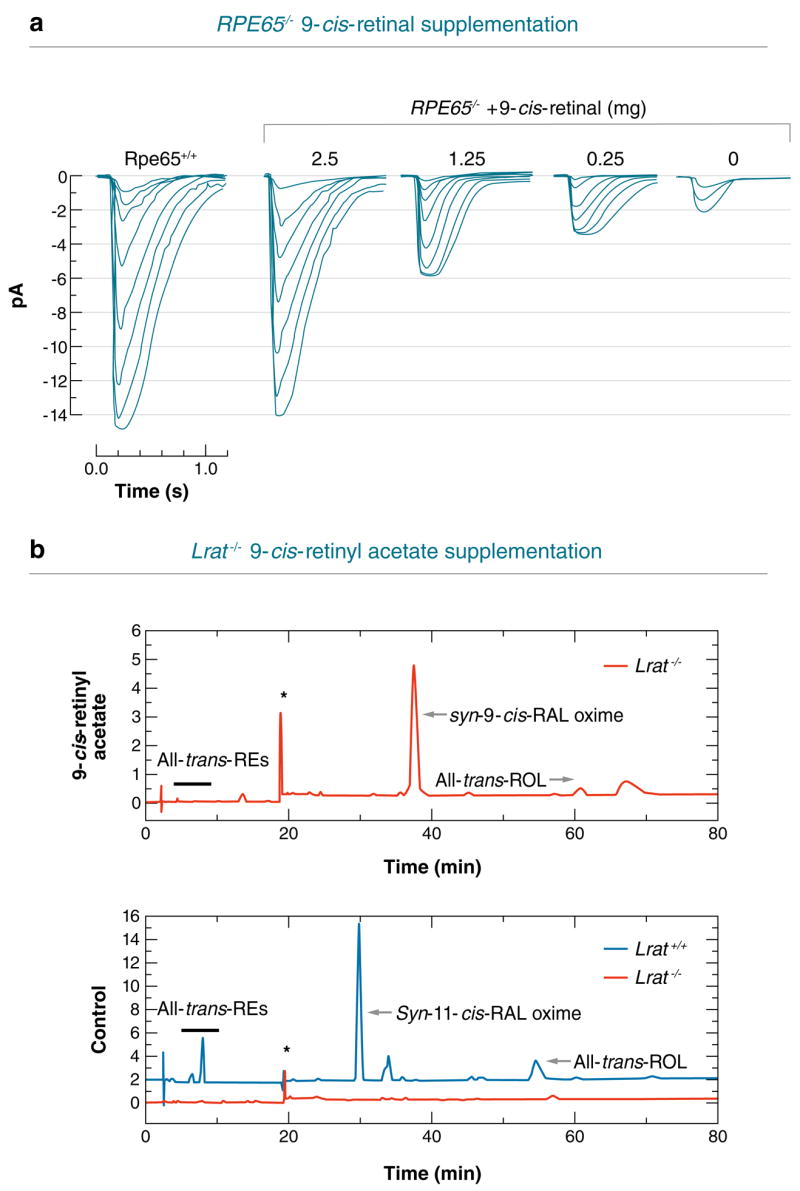
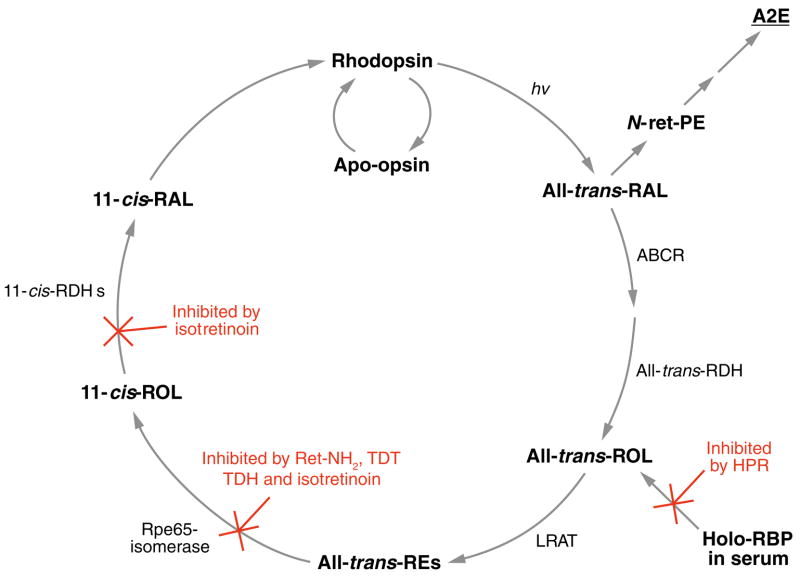
Absorption of a photon by an opsin pigment causes isomerization of the chromophore from 11-cis-retinaldehyde to all-trans-retinaldehyde. Regeneration of visual chromophore following light exposure is dependent on an enzyme pathway called the retinoid or visual cycle. Our understanding of this pathway has been greatly facilitated by the identification of disease-causing mutations in the genes coding for visual cycle enzymes. Defects in nearly every step of this pathway are responsible for human-inherited retinal dystrophies. These retinal dystrophies can be divided into two etiologic groups. One involves the impaired synthesis of visual chromophore. The second involves accumulation of cytotoxic products derived from all-trans-retinaldehyde. Gene therapy has been successfully used in animal models of these diseases to rescue the function of enzymes involved in chromophore regeneration, restoring vision. Dystrophies resulting from impaired chromophore synthesis can also be treated by supplementation with a chromophore analog. Dystrophies resulting from the accumulation of toxic pigments can be treated pharmacologically by inhibiting the visual cycle, or limiting the supply of vitamin A to the eyes. Recent progress in both areas provides hope that multiple inherited retinal diseases will soon be treated by pharmaceutical intervention.
Figures
References
LITERATURE CITED
-
- Okada T, Ernst OP, Palczewski K, Hofmann KP. Activation of rhodopsin: new insights from structural and biochemical studies. Trends Biochem Sci. 2001;26:318–24. - PubMed
-
- Arshavsky VY, Lamb TD, Pugh EN. G proteins and phototransduction. Annu Rev Physiol. 2002;64:153–87. - PubMed
-
- Rando RR, Bangerter FW. The rapid intermembraneous transfer of retinoids. Biochem Biophys Res Commun. 1982;104:430–36. - PubMed
-
- Groenendijk GW, De Grip WJ, Daemen FJ. Rod outer segment membranes: good acceptors for retinoids? Vision Res. 1984;24:1623–27. - PubMed
RELATED REVIEWS
-
- Mark M, Ghyselinck NB, Chambon P. Function of retinoid nuclear receptors: lessons from genetic and pharmacological dissections of the retinoic acid signaling pathway during mouse embryogenesis. Annu Rev Pharmacol Toxicol. 2006;46:451–80. - PubMed
-
- Chytil F. Rough and rocky road to the retinoid revolution. Annu Rev Nutr. 2003;23:1–16. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
