

A molecular mechanism for osmolyte-induced protein stability
- PMID: 16968772
- PMCID: PMC1564065
- DOI: 10.1073/pnas.0606236103
A molecular mechanism for osmolyte-induced protein stability
Erratum in
- Proc Natl Acad Sci U S A. 2006 Nov 7;103(45):17064
Abstract
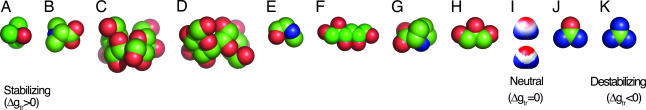
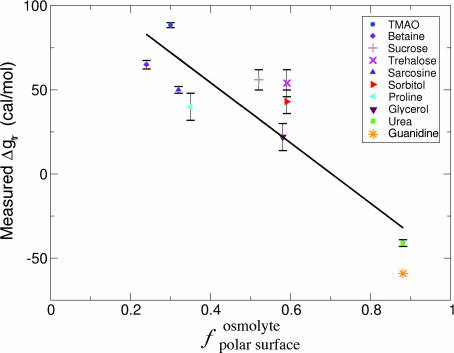
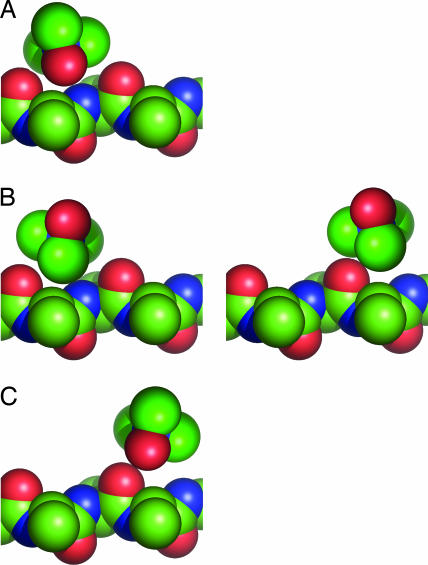
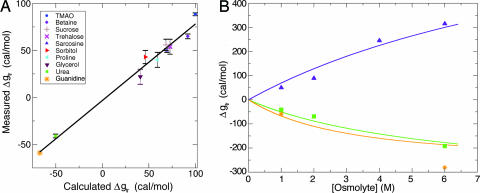
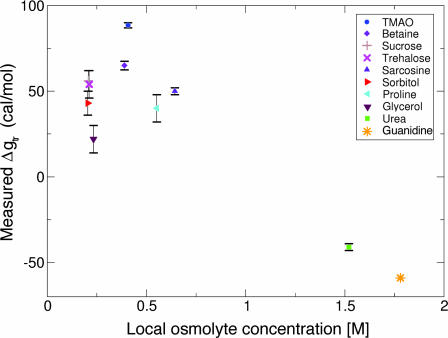
Osmolytes are small organic compounds that affect protein stability and are ubiquitous in living systems. In the equilibrium protein folding reaction, unfolded (U) native (N), protecting osmolytes push the equilibrium toward N, whereas denaturing osmolytes push the equilibrium toward U. As yet, there is no universal molecular theory that can explain the mechanism by which osmolytes interact with the protein to affect protein stability. Here, we lay the groundwork for such a theory, starting with a key observation: the transfer free energy of protein backbone from water to a water/osmolyte solution, Deltagtr, is negatively correlated with an osmolyte's fractional polar surface area. Deltagtr measures the degree to which an osmolyte stabilizes a protein. Consequently, a straightforward interpretation of this correlation implies that the interaction between the protein backbone and osmolyte polar groups is more favorable than the corresponding interaction with nonpolar groups. Such an interpretation immediately suggests the existence of a universal mechanism involving osmolyte, backbone, and water. We test this idea by using it to construct a quantitative solvation model in which backbone/solvent interaction energy is a function of interactant polarity, and the number of energetically equivalent ways of realizing a given interaction is a function of interactant surface area. Using this model, calculated Deltagtr values show a strong correlation with measured values (R = 0.99). In addition, the model correctly predicts that protecting/denaturing osmolytes will be preferentially excluded/accumulated around the protein backbone. Taken together, these model-based results rationalize the dominant interactions observed in experimental studies of osmolyte-induced protein stabilization and denaturation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
