

Antineoplastic effects of rosiglitazone and PPARgamma transactivation in neuroblastoma cells
- PMID: 16969347
- PMCID: PMC2360542
- DOI: 10.1038/sj.bjc.6603344
Antineoplastic effects of rosiglitazone and PPARgamma transactivation in neuroblastoma cells
Abstract
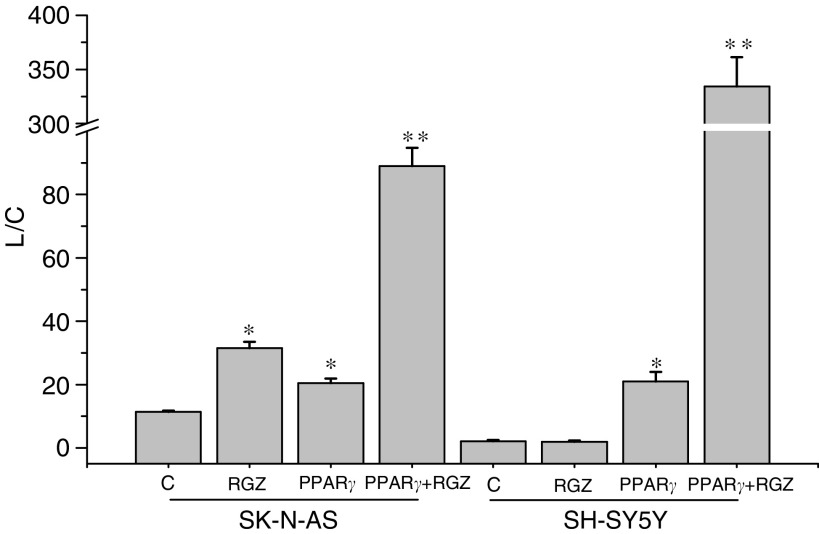
Neuroblastoma (NB) is the most common extracranial solid tumour in infants. Unfortunately, most children present with advanced disease and have a poor prognosis. In the present study, we evaluated the role of the peroxisome proliferator-activated receptor gamma (PPARgamma) agonist rosiglitazone (RGZ) in two NB cell lines (SK-N-AS and SH-SY5Y), which express PPARgamma. Rosiglitazone decreased cell proliferation and viability to a greater extent in SK-N-AS than in SH-SY5Y. Furthermore, 20 microM RGZ significantly inhibited cell adhesion, invasiveness and apoptosis in SK-N-AS, but not in SH-SY5Y. Because of the different response of SK-N-AS and SH-SY5Y cells to RGZ, the function of PPARgamma as a transcriptional activator was assessed. Noticeably, transient transcription experiments with a PPARgamma responsive element showed that RGZ induced a three-fold increase of the reporter activity in SK-N-AS, whereas no effect was observed in SH-SY5Y. The different PPARgamma activity may be likely due to the markedly lower amount of phopshorylated (i.e. inactive) protein observed in SK-N-AS. To our knowledge, this is the first demonstration that the differential response of NB cells to RGZ may be related to differences in PPARgamma transactivation. This finding indicates that PPARgamma activity may be useful to select those patients, for whom PPARgamma agonists may have a beneficial therapeutic effect.
Figures







References
-
- Benvenuti S, Luciani P, Vannelli GB, Gelmini S, Franceschi E, Serio M, Peri A (2005) Estrogen and selective estrogen receptor modulators exert neuroprotective effects and stimulate the expression of selective Alzheimer's disease indicator-1, a recently discovered antiapoptotic gene, in human neuroblast long-term cell cultures. J Clin Endocrinol Metab 90: 1775–1782 - PubMed
-
- Burstein HJ, Demetri GD, Mueller E, Sarraf P, Spiegelman BM, Winer EP (2003) Use of the peroxisome proliferator-activated receptor (PPAR) gamma ligand troglitazone as treatment for refractory breast cancer: a phase II study. Breast Cancer Res Treat 79: 391–397 - PubMed
-
- Carloni V, Romanelli RG, Mercurio AM, Pinzani M, Laffi G, Cotrozzi G, Gentilini P (1998) Knockout of alpha6 beta1-integrin expression reverses the transformed phenotype of hepatocarcinoma cells. Gastroenterology 115: 433–442 - PubMed
-
- Cavallaro U, Christofori G (2004) Multitasking in tumor progression: signaling functions of cell adhesion molecules. Ann NY Acad Sci 1014: 58–66 - PubMed
-
- Chantrain CF, Shimada H, Jodele S, Groshen S, Ye W, Shalinsky DR, Werb Z, Coussens LM, DeClerck YA (2004) Stromal matrix metalloproteinase-9 regulates the vascular architecture in neuroblastoma by promoting pericyte recruitment. Cancer Res 64: 1675–1686 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
