

Mechanically induced actin-mediated rocketing of phagosomes
- PMID: 16971511
- PMCID: PMC1635377
- DOI: 10.1091/mbc.e06-04-0365
Mechanically induced actin-mediated rocketing of phagosomes
Abstract
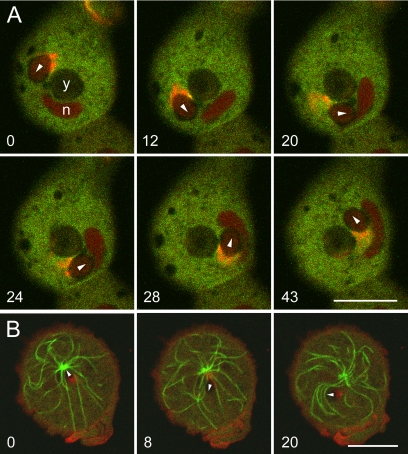
Actin polymerization can be induced in Dictyostelium by compressing the cells to bring phagosomes filled with large particles into contact with the plasma membrane. Asymmetric actin assembly results in rocketing movement of the phagosomes. We show that the compression-induced assembly of actin at the cytoplasmic face of the plasma membrane involves the Arp2/3 complex. We also identify two other proteins associated with the mechanically induced actin assembly. The class I myosin MyoB accumulates at the plasma membrane-phagosome interface early during the initiation of the response, and coronin is recruited as the actin filaments are disassembling. The forces generated by rocketing phagosomes are sufficient to push the entire microtubule apparatus forward and to dislocate the nucleus.
Figures






Similar articles
-
Dynamin A, Myosin IB and Abp1 couple phagosome maturation to F-actin binding.Traffic. 2012 Jan;13(1):120-30. doi: 10.1111/j.1600-0854.2011.01296.x. Epub 2011 Oct 26. Traffic. 2012. PMID: 22008230
-
Phagocyte meets prey: uptake, internalization, and killing of bacteria by Dictyostelium amoebae.Eur J Cell Biol. 2006 Sep;85(9-10):1001-10. doi: 10.1016/j.ejcb.2006.05.004. Epub 2006 Jun 16. Eur J Cell Biol. 2006. PMID: 16782228
-
Mutants in the Dictyostelium Arp2/3 complex and chemoattractant-induced actin polymerization.Exp Cell Res. 2007 Jul 15;313(12):2563-74. doi: 10.1016/j.yexcr.2007.04.029. Epub 2007 May 6. Exp Cell Res. 2007. PMID: 17553489
-
Role of microtubules and myosins in Fc gamma receptor-mediated phagocytosis.Front Biosci. 2006 May 1;11:1479-90. doi: 10.2741/1897. Front Biosci. 2006. PMID: 16368530 Review.
-
How to build a phagosome: new concepts for an old process.Curr Opin Cell Biol. 2018 Feb;50:57-63. doi: 10.1016/j.ceb.2018.01.009. Epub 2018 Feb 20. Curr Opin Cell Biol. 2018. PMID: 29471269 Review.
Cited by
-
Open access microfluidic device for the study of cell migration during chemotaxis.Integr Biol (Camb). 2010 Nov;2(11-12):648-58. doi: 10.1039/c0ib00110d. Epub 2010 Oct 15. Integr Biol (Camb). 2010. PMID: 20949221 Free PMC article.
-
Eukaryotic chemotaxis: a network of signaling pathways controls motility, directional sensing, and polarity.Annu Rev Biophys. 2010;39:265-89. doi: 10.1146/annurev.biophys.093008.131228. Annu Rev Biophys. 2010. PMID: 20192768 Free PMC article. Review.
-
Retrieval of the vacuolar H-ATPase from phagosomes revealed by live cell imaging.PLoS One. 2010 Jan 5;5(1):e8585. doi: 10.1371/journal.pone.0008585. PLoS One. 2010. PMID: 20052281 Free PMC article.
-
Dictyostelium myosin 1F and myosin 1E inhibit actin waves in a lipid-binding-dependent and motor-independent manner.Cytoskeleton (Hoboken). 2020 Aug;77(8):295-302. doi: 10.1002/cm.21627. Cytoskeleton (Hoboken). 2020. PMID: 32734648 Free PMC article.
-
Coronin 1B regulates S1P-induced human lung endothelial cell chemotaxis: role of PLD2, protein kinase C and Rac1 signal transduction.PLoS One. 2013 May 8;8(5):e63007. doi: 10.1371/journal.pone.0063007. Print 2013. PLoS One. 2013. PMID: 23667561 Free PMC article.
References
-
- Aubrey L., Klein G., Martiel J.-L., Satre M. Kinetics of endosomal pH evolution in Dictyostelium discoideum amoebae. Study by fluorescence spectroscopy. J. Cell Sci. 1993;105:861–866. - PubMed
-
- Axelrod D. Total internal reflection fluorescence microscopy in cell biology. Traffic. 2001;2:764–774. - PubMed
-
- Bernheim-Groswasser A., Wiesner S., Golsteyn R. M., Carlier M. F., Sykes C. The dynamics of actin-based motility depend on surface parameters. Nature. 2002;417:308–311. - PubMed
-
- Bretschneider T., Diez S., Anderson K., Heuser J., Clarke M., Müller-Taubenberger A., Köhler J., Gerisch G. Dynamic actin patterns and Arp2/3 assembly at the substrate-attached surface of motile cells. Curr. Biol. 2004;14:1–10. - PubMed
-
- Cai L., Holoweckyj N., Schaller M. D., Bear J. E. Phosphorylation of coronin 1B by protein kinase C regulates interaction with Arp2/3 and cell motility. J. Biol. Chem. 2005;280:31913–31923. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
