

Dynamic regulation of cAMP synthesis through anchored PKA-adenylyl cyclase V/VI complexes
- PMID: 16973443
- PMCID: PMC3941446
- DOI: 10.1016/j.molcel.2006.07.025
Dynamic regulation of cAMP synthesis through anchored PKA-adenylyl cyclase V/VI complexes
Abstract
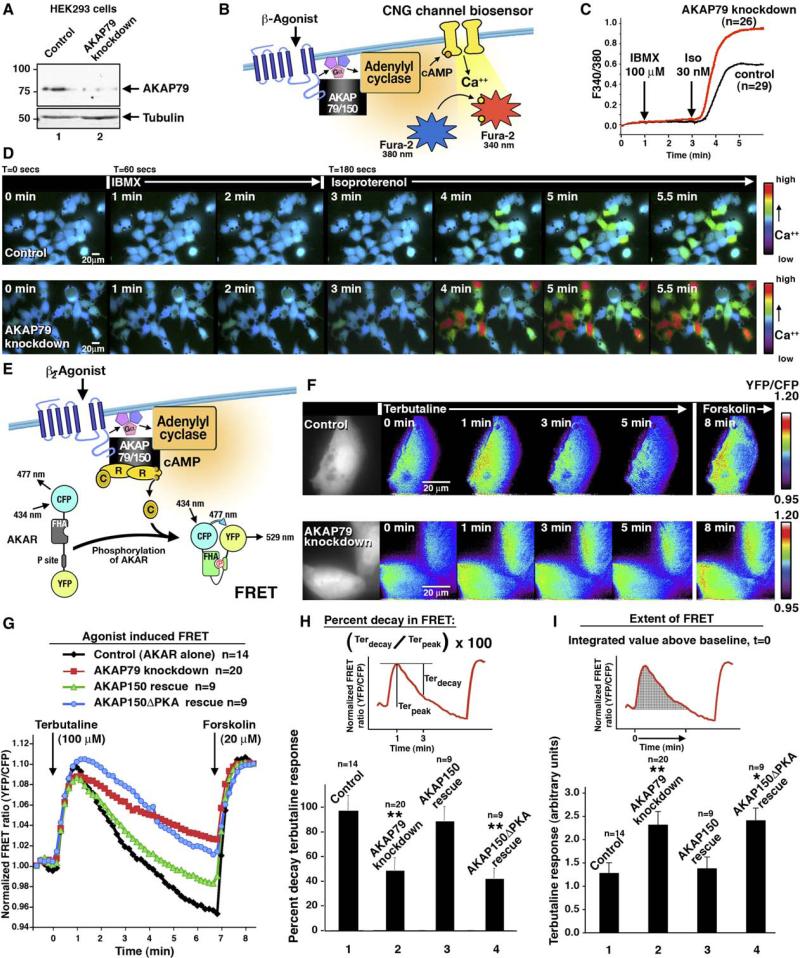
Spatiotemporal organization of cAMP signaling begins with the tight control of second messenger synthesis. In response to agonist stimulation of G protein-coupled receptors, membrane-associated adenylyl cyclases (ACs) generate cAMP that diffuses throughout the cell. The availability of cAMP activates various intracellular effectors, including protein kinase A (PKA). Specificity in PKA action is achieved by the localization of the enzyme near its substrates through association with A-kinase anchoring proteins (AKAPs). Here, we provide evidence for interactions between AKAP79/150 and ACV and ACVI. PKA anchoring facilitates the preferential phosphorylation of AC to inhibit cAMP synthesis. Real-time cellular imaging experiments show that PKA anchoring with the cAMP synthesis machinery ensures rapid termination of cAMP signaling upon activation of the kinase. This protein configuration permits the formation of a negative feedback loop that temporally regulates cAMP production.
Figures



References
-
- Asirvatham AL, Galligan SG, Schillace RV, Davey MP, Vasta V, Beavo JA, Carr DW. A-kinase anchoring proteins interact with phosphodiesterases in T lymphocyte cell lines. J. Immunol. 2004;173:4806–4814. - PubMed
-
- Baillie GS, Scott JD, Houslay MD. Compartmentalisation of phosphodiesterases and protein kinase A: opposites attract. FEBS Lett. 2005;579:3264–3270. - PubMed
-
- Bohn LM, Lefkowitz RJ, Gainetdinov RR, Peppel K, Caron MG, Lin FT. Enhanced morphine analgesia in mice lacking beta-arrestin 2. Science. 1999;286:2495–2498. - PubMed
-
- Bos JL. Epac: a new cAMP target and new avenues in cAMP research. Nat. Rev. Mol. Cell Biol. 2003;4:733–738. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
