

Essential functions of the unique N-terminal region of the varicella-zoster virus glycoprotein E ectodomain in viral replication and in the pathogenesis of skin infection
- PMID: 16973553
- PMCID: PMC1617235
- DOI: 10.1128/JVI.00533-06
Essential functions of the unique N-terminal region of the varicella-zoster virus glycoprotein E ectodomain in viral replication and in the pathogenesis of skin infection
Abstract
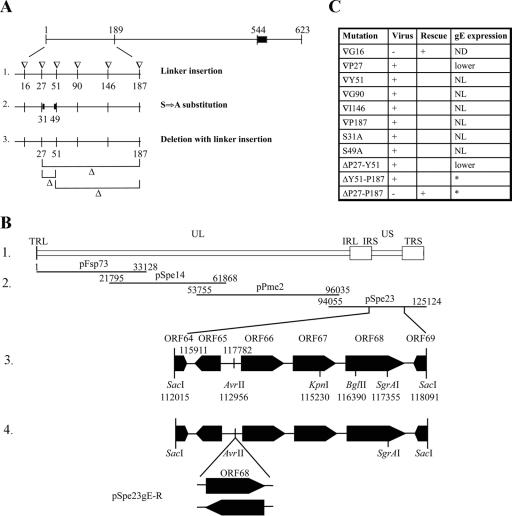
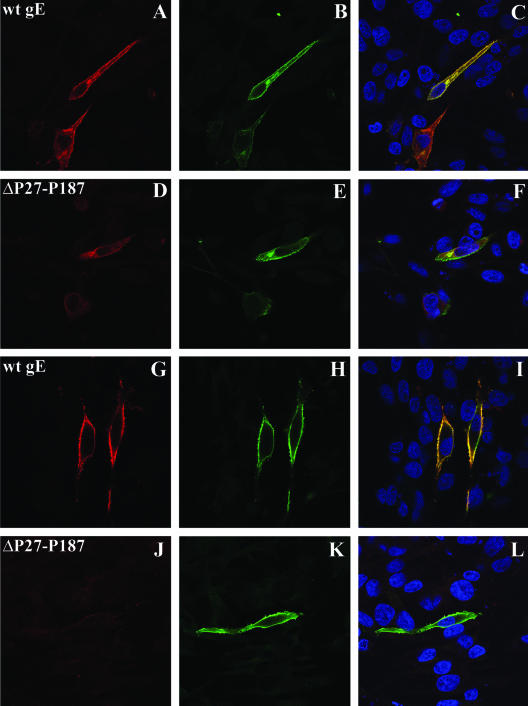
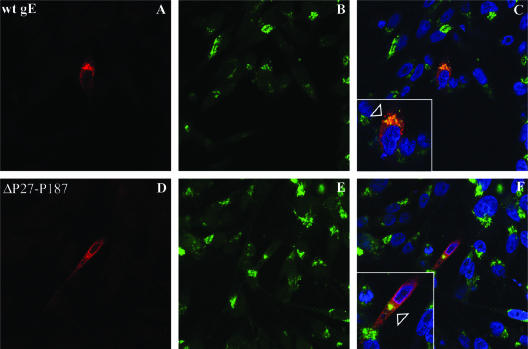
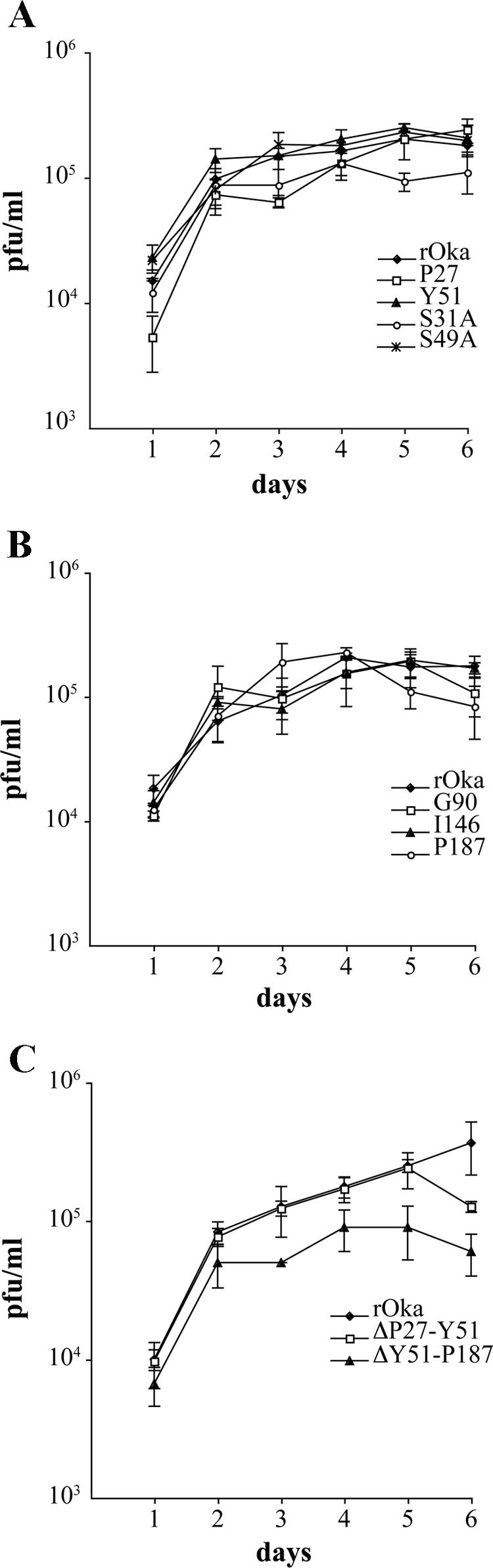
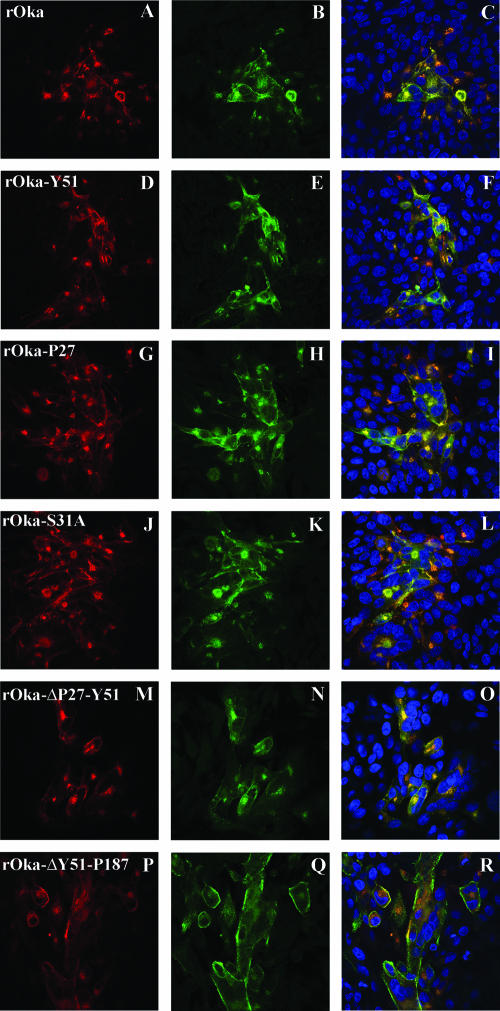
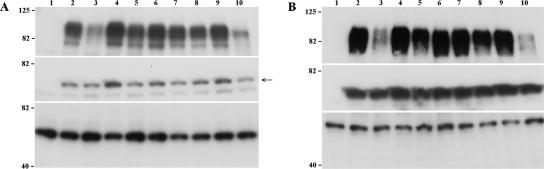
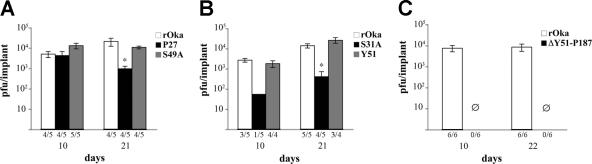
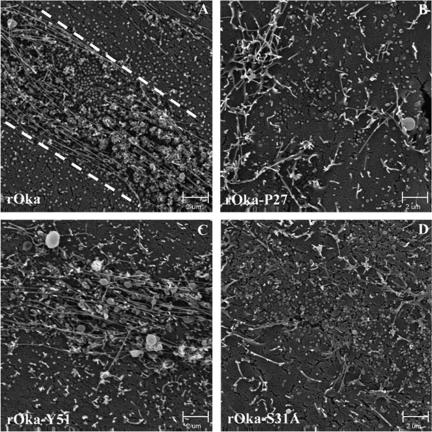
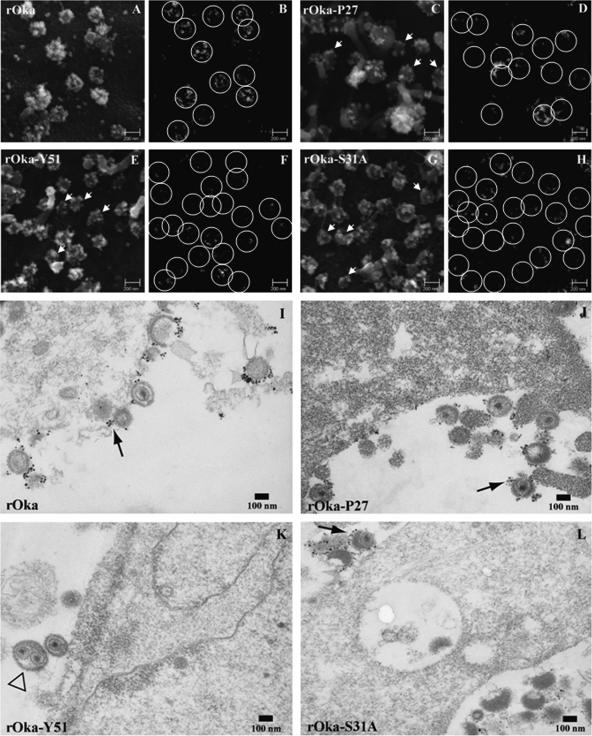
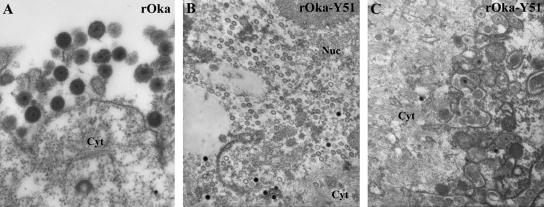
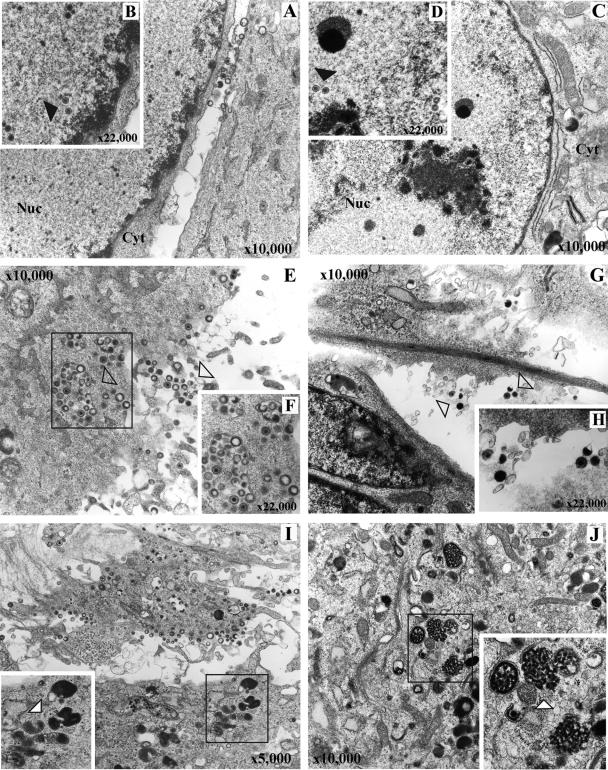
Varicella-zoster virus (VZV) glycoprotein E (gE) is a multifunctional protein important for cell-cell spread, envelopment, and possibly entry. In contrast to other alphaherpesviruses, gE is essential for VZV replication. Interestingly, the N-terminal region of gE, comprised of amino acids 1 to 188, was shown not to be conserved in the other alphaherpesviruses by bioinformatics analysis. Mutational analysis was performed to investigate the functions associated with this unique gE N-terminal region. Linker insertions, serine-to-alanine mutations, and deletions were introduced in the gE N-terminal region in the VZV genome, and the effects of these mutations on virus replication and cell-cell spread, gE trafficking and localization, virion formation, and replication in vivo in the skin were analyzed. In summary, mutagenesis of the gE N-terminal region identified a new functional region in the VZV gE ectodomain essential for cell-cell spread and the pathogenesis of VZV skin tropism and demonstrated that different subdomains of the unique N-terminal region had specific roles in viral replication, cell-cell spread, and secondary envelopment.
Figures

References
-
- Arvin, A. M. 2001. Varicella-zoster virus, p. 2731-2768. In D. M. Knipe, P. M. Howley, D. E. Griffin, R. A. Lamb, M. A. Martin, B. Roizman, and S. E. Straus (ed.), Fields virology, 4th ed., vol. 2. Lippincott Williams & Wilkins, Philadelphia, Pa.
-
- Baiker, A., C. Bagowski, H. Ito, M. Sommer, L. Zerboni, K. Fabel, J. Hay, W. Ruyechan, and A. M. Arvin. 2004. The immediate-early 63 protein of varicella-zoster virus: analysis of functional domains required for replication in vitro and for T-cell and skin tropism in the SCIDhu model in vivo. J. Virol. 78:1181-1194. - PMC - PubMed
-
- Bailey, T. L., and C. Elkan. 1994. Fitting a mixture model by expectation maximization to discover motifs in biopolymers. Proc. Int. Conf. Intell. Syst. Mol. Biol. 2:28-36. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
