

Mobilization of full-length Semliki Forest virus replicon by retrovirus particles
- PMID: 16973593
- PMCID: PMC1617220
- DOI: 10.1128/JVI.00664-06
Mobilization of full-length Semliki Forest virus replicon by retrovirus particles
Abstract
Conciliating biosafety with efficient gene transfer remains a constant concern in the development of retroviral vectors. Semliki Forest virus (SFV) replicons allow important retroviral vector production with interesting features. It is noteworthy that retroviruses have the ability to package Psi+ and, to some extent, Psi- cellular RNAs. Therefore, it was important to study the retroviral transfer of highly abundant SFV genomes expressing retroviral proteins. Here, we show that full-length SFV-vector replicons, with or without Psi, are efficiently packaged into retrovirus particles. Mechanistically, our data suggest that SFV packaging is the sum of its retroviral nucleocapsid-dependent recruitment together with a passive hijacking of membrane-anchored SFV replicon. A direct consequence of this phenomenon is the formation of particles harboring autonomous replicative abilities and contaminating vector preparations. Importantly, we confirm that retroviral SFV mobilization is not an exclusive feature of murine gamma retroviruses, since it is also observed using lentivectors.
Figures
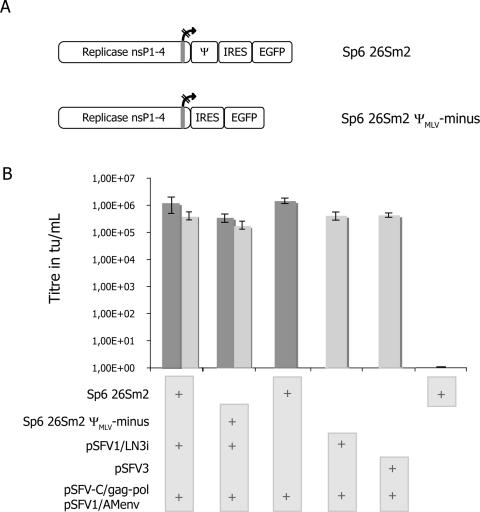
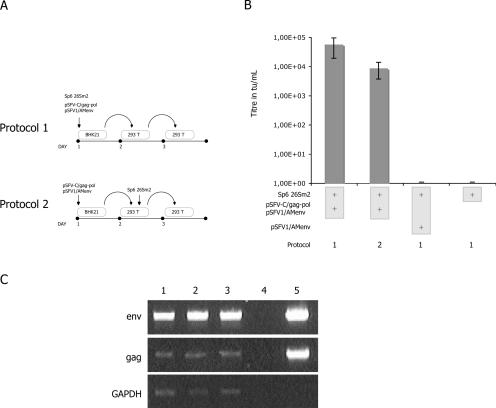
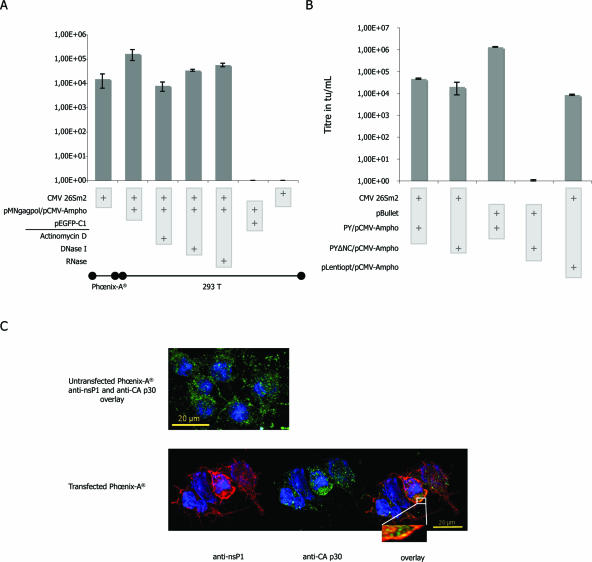
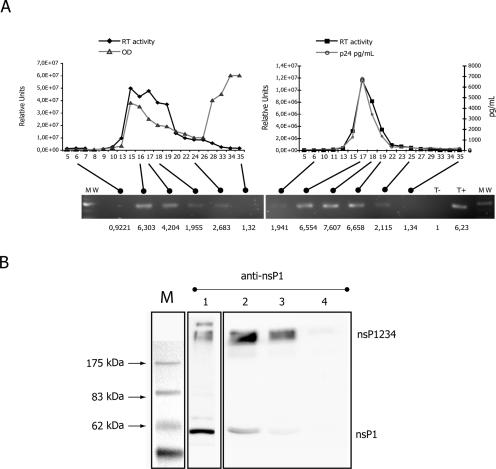
References
-
- Berglund, P., M. Sjoberg, H. Garoff, G. J. Atkins, B. J. Sheahan, and P. Liljestrom. 1993. Semliki Forest virus expression system: production of conditionally infectious recombinant particles. Biotechnology (NY) 11:916-920. - PubMed
-
- Bushman, F. 2002. Targeting retroviral integration? Mol. Ther. 6:570-571. - PubMed
-
- Cornetta, K., R. A. Morgan, and W. F. Anderson. 1991. Safety issues related to retroviral mediated gene transfer in humans. Hum. Gene Ther. 2:5-14. - PubMed
-
- Diatta, A., E. Piver, C. Collin, P. Vaudin, and J. C. Pages. 2005. Semliki Forest virus-derived virus-like particles: characterization of their production and transduction pathways. J. Gen. Virol. 86:3129-3136. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
