

CRACM1 multimers form the ion-selective pore of the CRAC channel
- PMID: 16978865
- PMCID: PMC5685803
- DOI: 10.1016/j.cub.2006.08.085
CRACM1 multimers form the ion-selective pore of the CRAC channel
Abstract
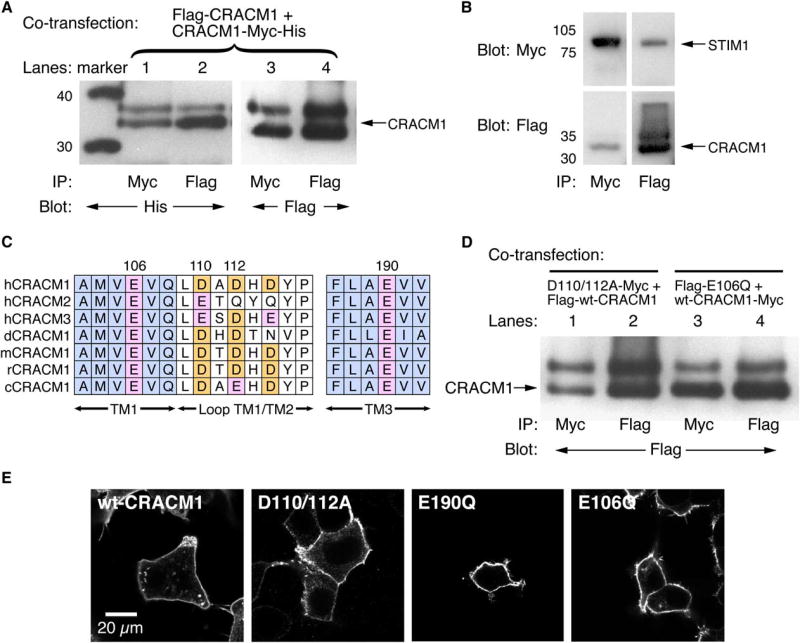
Receptor-mediated Ca(2+) release from the endoplasmic reticulum (ER) is often followed by Ca(2+) entry through Ca(2+)-release-activated Ca(2+) (CRAC) channels in the plasma membrane . RNAi screens have identified STIM1 as the putative ER Ca(2+) sensor and CRACM1 (Orai1; ) as the putative store-operated Ca(2+) channel. Overexpression of both proteins is required to reconstitute CRAC currents (I(CRAC); ). We show here that CRACM1 forms multimeric assemblies that bind STIM1 and that acidic residues in the transmembrane (TM) and extracellular domains of CRACM1 contribute to the ionic selectivity of the CRAC-channel pore. Replacement of the conserved glutamate in position 106 of the first TM domain of CRACM1 with glutamine (E106Q) acts as a dominant-negative protein, and substitution with aspartate (E106D) enhances Na(+), Ba(2+), and Sr(2+) permeation relative to Ca(2+). Mutating E190Q in TM3 also affects channel selectivity, suggesting that glutamate residues in both TM1 and TM3 face the lumen of the pore. Furthermore, mutating a putative Ca(2+) binding site in the first extracellular loop of CRACM1 (D110/112A) enhances monovalent cation permeation, suggesting that these residues too contribute to the coordination of Ca(2+) ions to the pore. Our data provide unequivocal evidence that CRACM1 multimers form the Ca(2+)-selective CRAC-channel pore.
Figures



References
-
- Hoth M, Penner R. Depletion of intracellular calcium stores activates a calcium current in mast cells. Nature. 1992;355:353–356. - PubMed
-
- Parekh AB, Penner R. Store depletion and calcium influx. Physiol. Rev. 1997;77:901–930. - PubMed
-
- Parekh AB, Putney JW., Jr Store-operated calcium channels. Physiol. Rev. 2005;85:757–810. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
