

Distinct functions of dispersed GATA factor complexes at an endogenous gene locus
- PMID: 16980610
- PMCID: PMC1592882
- DOI: 10.1128/MCB.01033-06
Distinct functions of dispersed GATA factor complexes at an endogenous gene locus
Abstract
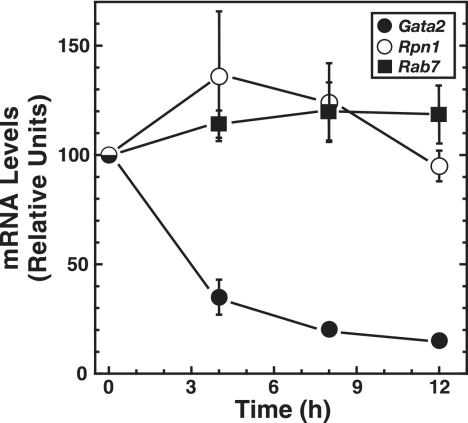
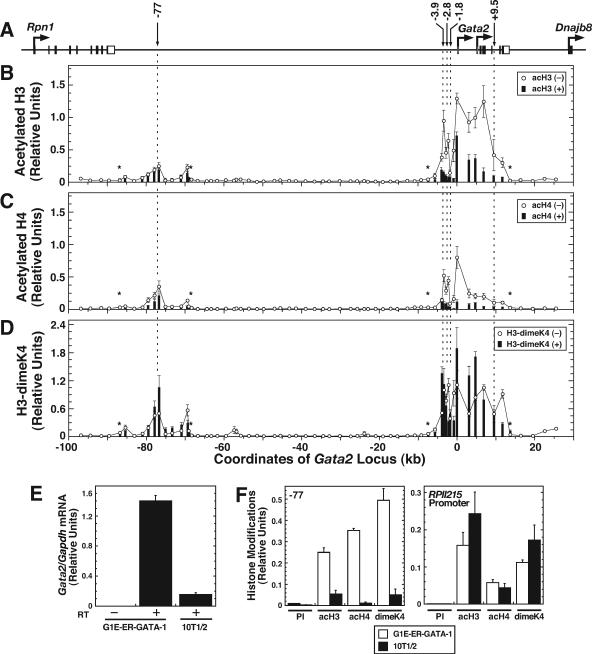
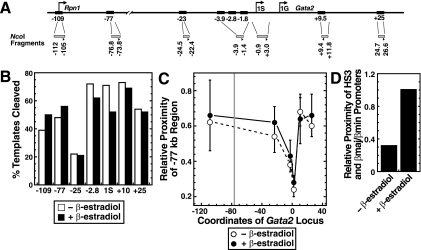
The reciprocal expression of GATA-1 and GATA-2 during hematopoiesis is an important determinant of red blood cell development. Whereas Gata2 is preferentially transcribed early in hematopoiesis, elevated GATA-1 levels result in GATA-1 occupancy at sites upstream of the Gata2 locus and transcriptional repression. GATA-2 occupies these sites in the transcriptionally active locus, suggesting that a "GATA switch" abrogates GATA-2-mediated positive autoregulation. Chromatin immunoprecipitation (ChIP) coupled with genomic microarray analysis and quantitative ChIP analysis with GATA-1-null cells expressing an estrogen receptor ligand binding domain fusion to GATA-1 revealed additional GATA switches 77 kb upstream of Gata2 and within intron 4 at +9.5 kb. Despite indistinguishable GATA-1 occupancy at -77 kb and +9.5 kb versus other GATA switch sites, GATA-1 functioned uniquely at the different regions. GATA-1 induced histone deacetylation at and near Gata2 but not at the -77 kb region. The -77 kb region, which was DNase I hypersensitive in both active and inactive states, conferred equivalent enhancer activities in GATA-1- and GATA-2-expressing cells. By contrast, the +9.5 kb region exhibited considerably stronger enhancer activity in GATA-2- than in GATA-1-expressing cells, and other GATA switch sites were active only in GATA-1- or GATA-2-expressing cells. Chromosome conformation capture analysis demonstrated higher-order interactions between the -77 kb region and Gata2 in the active and repressed states. These results indicate that dispersed GATA factor complexes function via long-range chromatin interactions and qualitatively distinct activities to regulate Gata2 transcription.
Figures






Similar articles
-
Context-dependent GATA factor function: combinatorial requirements for transcriptional control in hematopoietic and endothelial cells.J Biol Chem. 2007 May 11;282(19):14665-74. doi: 10.1074/jbc.M700792200. Epub 2007 Mar 7. J Biol Chem. 2007. PMID: 17347142
-
GATA-1-mediated transcriptional repression yields persistent transcription factor IIB-chromatin complexes.J Biol Chem. 2006 Dec 8;281(49):37345-52. doi: 10.1074/jbc.M605774200. Epub 2006 Sep 8. J Biol Chem. 2006. PMID: 16963445
-
Dynamic GATA factor interplay at a multicomponent regulatory region of the GATA-2 locus.J Biol Chem. 2005 Jan 21;280(3):1724-32. doi: 10.1074/jbc.M406038200. Epub 2004 Oct 19. J Biol Chem. 2005. PMID: 15494394
-
GATA switches as developmental drivers.J Biol Chem. 2010 Oct 8;285(41):31087-93. doi: 10.1074/jbc.R110.159079. Epub 2010 Jul 29. J Biol Chem. 2010. PMID: 20670937 Free PMC article. Review.
-
GATA-related hematologic disorders.Exp Hematol. 2016 Aug;44(8):696-705. doi: 10.1016/j.exphem.2016.05.010. Epub 2016 May 25. Exp Hematol. 2016. PMID: 27235756 Review.
Cited by
-
Transcriptional mechanisms underlying hemoglobin synthesis.Cold Spring Harb Perspect Med. 2013 Sep 1;3(9):a015412. doi: 10.1101/cshperspect.a015412. Cold Spring Harb Perspect Med. 2013. PMID: 23838521 Free PMC article. Review.
-
Embryonic erythropoiesis and hemoglobin switching require transcriptional repressor ETO2 to modulate chromatin organization.Nucleic Acids Res. 2020 Oct 9;48(18):10226-10240. doi: 10.1093/nar/gkaa736. Nucleic Acids Res. 2020. PMID: 32960220 Free PMC article.
-
GATA2 deficiency elevates interferon regulatory factor-8 to subvert a progenitor cell differentiation program.Blood Adv. 2022 Mar 8;6(5):1464-1473. doi: 10.1182/bloodadvances.2021006182. Blood Adv. 2022. PMID: 35008108 Free PMC article.
-
Induction of hematopoietic and endothelial cell program orchestrated by ETS transcription factor ER71/ETV2.EMBO Rep. 2015 May;16(5):654-69. doi: 10.15252/embr.201439939. Epub 2015 Mar 23. EMBO Rep. 2015. PMID: 25802403 Free PMC article.
-
GATA2 +9.5 enhancer: from principles of hematopoiesis to genetic diagnosis in precision medicine.Curr Opin Hematol. 2020 May;27(3):163-171. doi: 10.1097/MOH.0000000000000576. Curr Opin Hematol. 2020. PMID: 32205587 Free PMC article. Review.
References
-
- Bresnick, E. H., K. D. Johnson, S.-I. Kim, and H. Im. 2006. Establishment and regulation of chromatin domains: mechanistic insights from studies of hemoglobin synthesis. Prog. Nucleic Acids Res. Mol. Biol. 81:435-471. - PubMed
-
- Bresnick, E. H., M. L. Martowicz, S. Pal, and K. D. Johnson. 2005. Developmental control via GATA factor interplay at chromatin domains. J. Cell. Physiol. 205:1-9. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources