

Persistence of cooperatively stabilized signaling clusters drives T-cell activation
- PMID: 16980618
- PMCID: PMC1592897
- DOI: 10.1128/MCB.00507-06
Persistence of cooperatively stabilized signaling clusters drives T-cell activation
Abstract
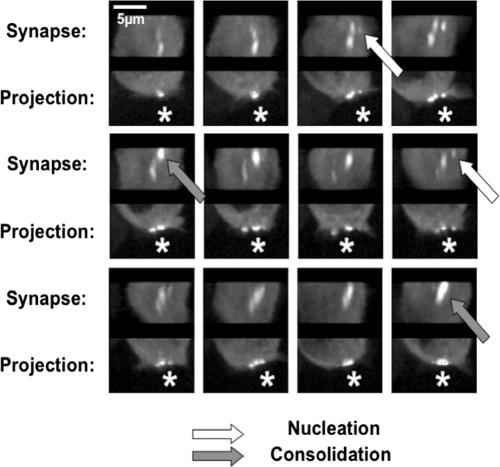
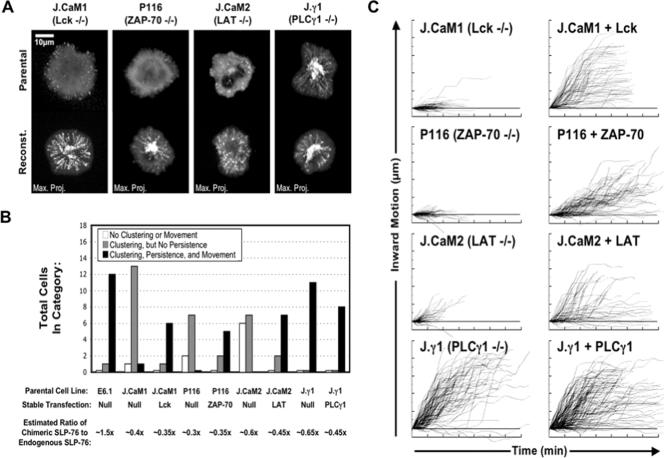
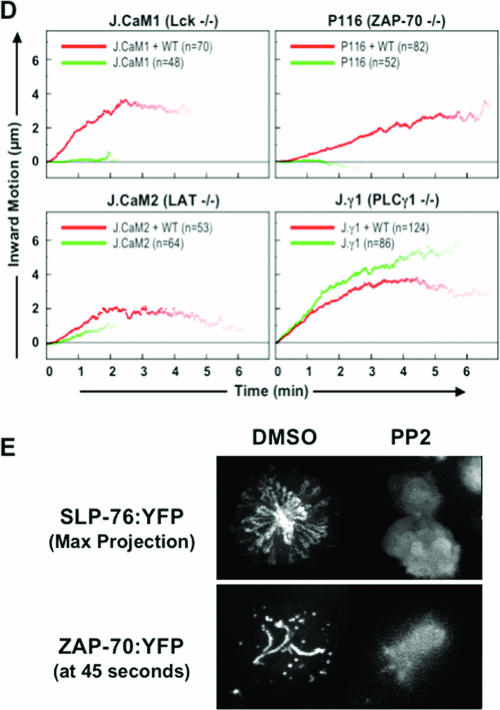
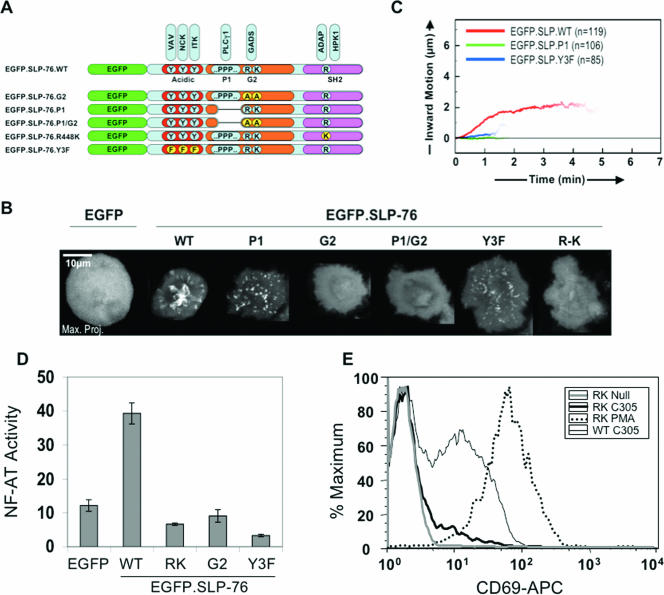
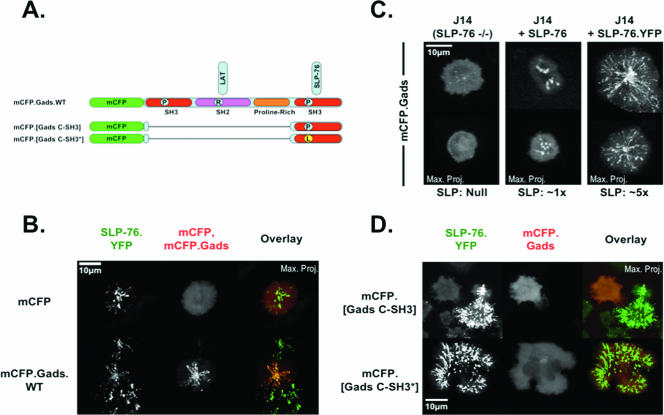
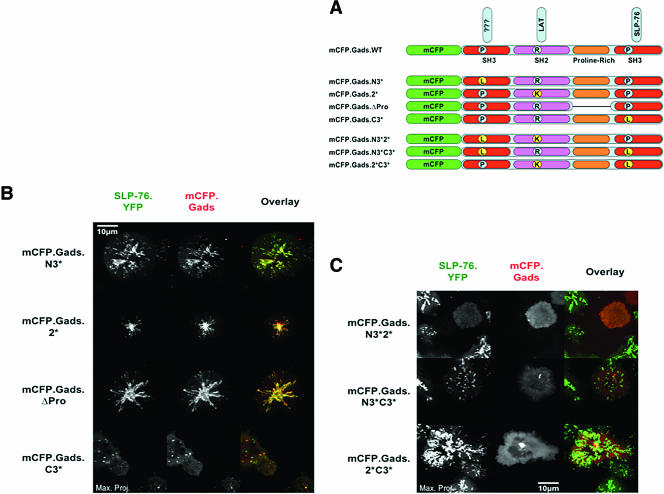
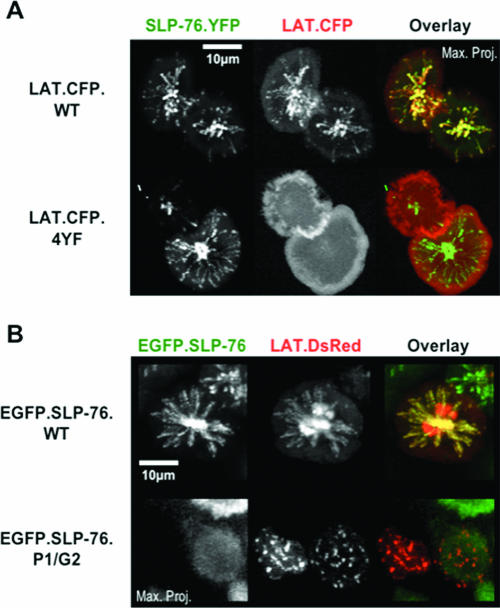
Antigen recognition triggers the recruitment of the critical adaptor protein SLP-76 to small macromolecular clusters nucleated by the T-cell receptor (TCR). These structures develop rapidly, in parallel with TCR-induced increases in tyrosine phosphorylation and cytosolic calcium, and are likely to contribute to TCR-proximal signaling. Previously, we demonstrated that these SLP-76-containing clusters segregate from the TCR and move towards the center of the contact interface. Neither the function of these clusters nor the structural requirements governing their persistence have been examined extensively. Here we demonstrate that defects in cluster assembly and persistence are associated with defects in T-cell activation in the absence of Lck, ZAP-70, or LAT. Clusters persist normally in the absence of phospholipase C-gamma1, indicating that in the absence of a critical effector, these structures are insufficient to drive T-cell activation. Furthermore, we show that the critical adaptors LAT and Gads localize with SLP-76 in persistent clusters. Mutational analyses of LAT, Gads, and SLP-76 indicated that multiple domains within each of these proteins contribute to cluster persistence. These data indicate that multivalent cooperative interactions stabilize these persistent signaling clusters, which may correspond to the functional complexes predicted by kinetic proofreading models of T-cell activation.
Figures
Similar articles
-
SH2 domain containing leukocyte phosphoprotein of 76-kDa (SLP-76) feedback regulation of ZAP-70 microclustering.Proc Natl Acad Sci U S A. 2010 Jun 1;107(22):10166-71. doi: 10.1073/pnas.0909112107. Epub 2010 May 13. Proc Natl Acad Sci U S A. 2010. PMID: 20534575 Free PMC article.
-
T cell activation at the immunological synapse: vesicles emerge for LATer signaling.Sci Signal. 2010 May 11;3(121):pe16. doi: 10.1126/scisignal.3121pe16. Sci Signal. 2010. PMID: 20460646 Review.
-
Recruitment of SLP-76 to the membrane and glycolipid-enriched membrane microdomains replaces the requirement for linker for activation of T cells in T cell receptor signaling.J Exp Med. 2000 Oct 2;192(7):1047-58. doi: 10.1084/jem.192.7.1047. J Exp Med. 2000. PMID: 11015445 Free PMC article.
-
Identification of a phospholipase C-gamma1 (PLC-gamma1) SH3 domain-binding site in SLP-76 required for T-cell receptor-mediated activation of PLC-gamma1 and NFAT.Mol Cell Biol. 2001 Jul;21(13):4208-18. doi: 10.1128/MCB.21.13.4208-4218.2001. Mol Cell Biol. 2001. PMID: 11390650 Free PMC article.
-
LAT, the linker for activation of T cells: a bridge between T cell-specific and general signaling pathways.Sci STKE. 2000 Dec 19;2000(63):re1. doi: 10.1126/stke.2000.63.re1. Sci STKE. 2000. PMID: 11752630 Review.
Cited by
-
Modeling and simulation of aggregation of membrane protein LAT with molecular variability in the number of binding sites for cytosolic Grb2-SOS1-Grb2.PLoS One. 2012;7(3):e28758. doi: 10.1371/journal.pone.0028758. Epub 2012 Mar 1. PLoS One. 2012. PMID: 22396725 Free PMC article.
-
Vitamin E reverses impaired linker for activation of T cells activation in T cells from aged C57BL/6 mice.J Nutr. 2009 Jun;139(6):1192-7. doi: 10.3945/jn.108.103416. Epub 2009 Apr 29. J Nutr. 2009. PMID: 19403707 Free PMC article.
-
A quantitative assessment of costimulation and phosphatase activity on microclusters in early T cell signaling.PLoS One. 2013 Oct 30;8(10):e79277. doi: 10.1371/journal.pone.0079277. eCollection 2013. PLoS One. 2013. PMID: 24205378 Free PMC article.
-
The Clinical Aspect of Adaptor Molecules in T Cell Signaling: Lessons Learnt From Inborn Errors of Immunity.Front Immunol. 2021 Aug 12;12:701704. doi: 10.3389/fimmu.2021.701704. eCollection 2021. Front Immunol. 2021. PMID: 34456914 Free PMC article. Review.
-
Keeping the (kinase) party going: SLP-76 and ITK dance to the beat.Sci STKE. 2007 Jul 24;2007(396):pe39. doi: 10.1126/stke.3962007pe39. Sci STKE. 2007. PMID: 17652306 Free PMC article. Review.
References
-
- Balamuth, F., D. Leitenberg, J. Unternaehrer, I. Mellman, and K. Bottomly. 2001. Distinct patterns of membrane microdomain partitioning in Th1 and Th2 cells. Immunity 15:729-738. - PubMed
-
- Barda-Saad, M., A. Braiman, R. Titerence, S. C. Bunnell, V. A. Barr, and L. E. Samelson. 2005. Dynamic molecular interactions linking the T cell antigen receptor to the actin cytoskeleton. Nat. Immunol. 6:80-89. - PubMed
-
- Berry, D. M., P. Nash, S. K. Liu, T. Pawson, and C. J. McGlade. 2002. A high-affinity Arg-X-X-Lys SH3 binding motif confers specificity for the interaction between Gads and SLP-76 in T cell signaling. Curr. Biol. 12:1336-1341. - PubMed
-
- Boerth, N. J., J. J. Sadler, D. E. Bauer, J. L. Clements, S. M. Gheith, and G. A. Koretzky. 2000. Recruitment of SLP-76 to the membrane and glycolipid-enriched membrane microdomains replaces the requirement for linker for activation of T cells in T cell receptor signaling. J. Exp. Med. 192:1047-1058. - PMC - PubMed
-
- Boniface, J. J., J. D. Rabinowitz, C. Wulfing, J. Hampl, Z. Reich, J. D. Altman, R. M. Kantor, C. Beeson, H. M. McConnell, and M. M. Davis. 1998. Initiation of signal transduction through the T cell receptor requires the multivalent engagement of peptide/MHC ligands. Immunity 9:459-466. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous