

MDM2 is required for suppression of apoptosis by activated Akt1 in salivary acinar cells
- PMID: 16982679
- PMCID: PMC1636839
- DOI: 10.1128/MCB.01846-05
MDM2 is required for suppression of apoptosis by activated Akt1 in salivary acinar cells
Abstract
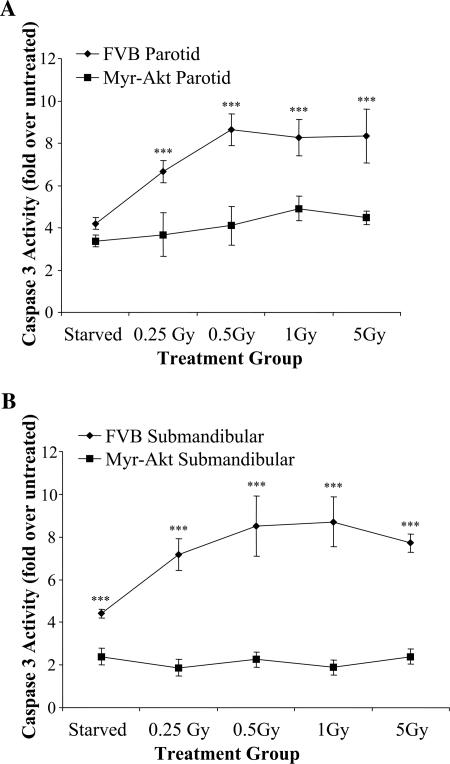
Chronic damage to the salivary glands is a common side effect following head and neck irradiation. It is hypothesized that irreversible damage to the salivary glands occurs immediately after radiation; however, previous studies with rat models have not shown a causal role for apoptosis in radiation-induced injury. We report that etoposide and gamma irradiation induce apoptosis of salivary acinar cells from FVB control mice in vitro and in vivo; however, apoptosis is reduced in transgenic mice expressing a constitutively activated mutant of Akt1 (myr-Akt1). Expression of myr-Akt1 in the salivary glands results in a significant reduction in phosphorylation of p53 at serine(18), total p53 protein accumulation, and p21(WAF1) or Bax mRNA following etoposide or gamma irradiation of primary salivary acinar cells. The reduced level of p53 protein in myr-Akt1 salivary glands corresponds with an increase in MDM2 phosphorylation in vivo, suggesting that the Akt/MDM2/p53 pathway is responsible for suppression of apoptosis. Dominant-negative Akt blocked phosphorylation of MDM2 in salivary acinar cells from myr-Akt1 transgenic mice. Reduction of MDM2 levels in myr-Akt1 primary salivary acinar cells with small interfering RNA increases the levels of p53 protein and renders these cells susceptible to etoposide-induced apoptosis in spite of the presence of activated Akt1. These results indicate that MDM2 is a critical substrate of activated Akt1 in the suppression of p53-dependent apoptosis in vivo.
Figures








Similar articles
-
Radiation-induced salivary gland dysfunction results from p53-dependent apoptosis.Int J Radiat Oncol Biol Phys. 2009 Feb 1;73(2):523-9. doi: 10.1016/j.ijrobp.2008.09.036. Int J Radiat Oncol Biol Phys. 2009. PMID: 19147016 Free PMC article.
-
Suppression of radiation-induced salivary gland dysfunction by IGF-1.PLoS One. 2009;4(3):e4663. doi: 10.1371/journal.pone.0004663. Epub 2009 Mar 2. PLoS One. 2009. PMID: 19252741 Free PMC article.
-
Insulin-like growth factor-1 preserves salivary gland function after fractionated radiation.Int J Radiat Oncol Biol Phys. 2010 Oct 1;78(2):579-86. doi: 10.1016/j.ijrobp.2010.03.035. Epub 2010 Jul 16. Int J Radiat Oncol Biol Phys. 2010. PMID: 20638195 Free PMC article.
-
Dose-response transition from cell cycle arrest to apoptosis with selective degradation of Mdm2 and p21WAF1/CIP1 in response to the novel anticancer agent, aminoflavone (NSC 686,288).Oncogene. 2007 Jul 19;26(33):4806-16. doi: 10.1038/sj.onc.1210283. Epub 2007 Feb 12. Oncogene. 2007. PMID: 17297446
-
Synergistic suppression of apoptosis in salivary acinar cells by IGF1 and EGF.Cell Death Differ. 2003 Mar;10(3):345-55. doi: 10.1038/sj.cdd.4401153. Cell Death Differ. 2003. PMID: 12700634 Free PMC article.
Cited by
-
Short-term and bystander effects of radiation on murine submandibular glands.Dis Model Mech. 2022 Nov 1;15(11):dmm049570. doi: 10.1242/dmm.049570. Epub 2022 Nov 10. Dis Model Mech. 2022. PMID: 36263624 Free PMC article.
-
Salivary gland function in nasopharyngeal carcinoma before and late after intensity-modulated radiotherapy evaluated by dynamic diffusion-weighted MR imaging with gustatory stimulation.BMC Oral Health. 2019 Dec 21;19(1):288. doi: 10.1186/s12903-019-0951-x. BMC Oral Health. 2019. PMID: 31864328 Free PMC article.
-
Administration of growth factors promotes salisphere formation from irradiated parotid salivary glands.PLoS One. 2018 Mar 28;13(3):e0193942. doi: 10.1371/journal.pone.0193942. eCollection 2018. PLoS One. 2018. PMID: 29590144 Free PMC article.
-
A single dose of radiation elicits comparable acute salivary gland injury to fractionated radiation.Dis Model Mech. 2024 Aug 1;17(8):dmm050733. doi: 10.1242/dmm.050733. Epub 2024 Aug 22. Dis Model Mech. 2024. PMID: 39086326 Free PMC article.
-
Radiation-induced changes in energy metabolism result in mitochondrial dysfunction in salivary glands.Sci Rep. 2024 Jan 8;14(1):845. doi: 10.1038/s41598-023-50877-9. Sci Rep. 2024. PMID: 38191641 Free PMC article.
References
-
- Abok, K., U. Brunk, B. Jung, and J. Ericsson. 1984. Morphologic and histochemical studies on the differing radiosensitivity of ductular and acinar cells of the rat submandibular gland. Virchows Arch. B Cell Pathol. Incl. Mol. Pathol. 45:443-460. - PubMed
-
- Alarcon-Vargas, D., and Z. Ronai. 2002. p53-Mdm2—the affair that never ends. Carcinogenesis 23:541-547. - PubMed
-
- Anderson, S. M., M. E. Reyland, S. Hunter, L. M. Deisher, K. A. Barzen, and D. O. Quissell. 1999. Etoposide-induced activation of c-jun N-terminal kinase (JNK) correlates with drug-induced apoptosis in salivary gland acinar cells. Cell Death Differ. 6:454-462. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous