

Ubiquitin-binding motifs in REV1 protein are required for its role in the tolerance of DNA damage
- PMID: 16982685
- PMCID: PMC1636806
- DOI: 10.1128/MCB.01118-06
Ubiquitin-binding motifs in REV1 protein are required for its role in the tolerance of DNA damage
Abstract
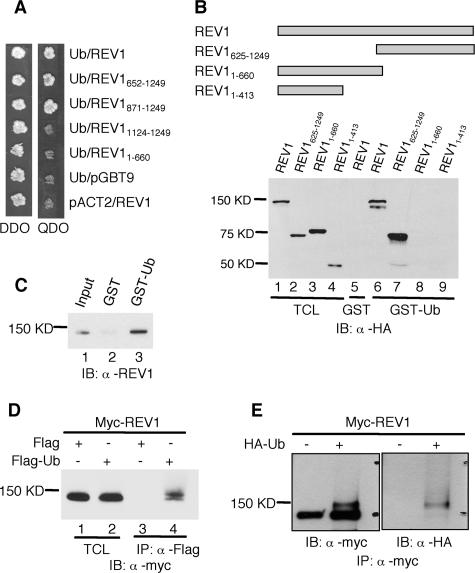
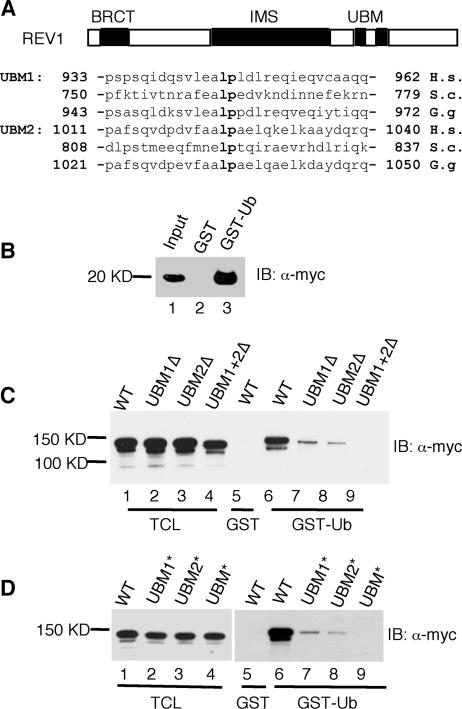
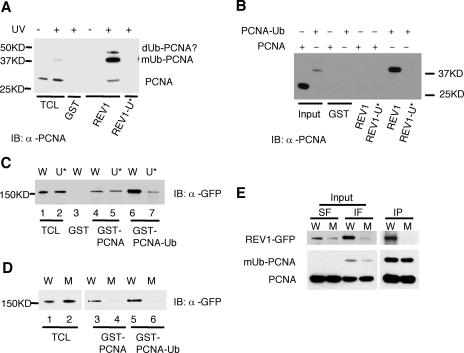
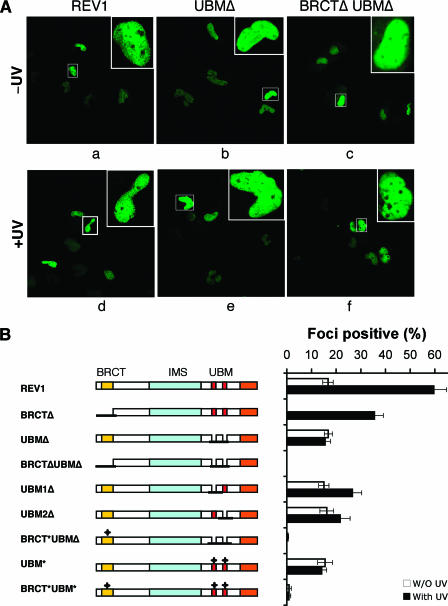
REV1 protein is a eukaryotic member of the Y family of DNA polymerases involved in the tolerance of DNA damage by replicative bypass. The precise role(s) of REV1 in this process is not known. Here we show, by using the yeast two-hybrid assay and the glutathione S-transferase pull-down assay, that mouse REV1 can physically interact with ubiquitin. The association of REV1 with ubiquitin requires the ubiquitin-binding motifs (UBMs) located at the C terminus of REV1. The UBMs also mediate the enhanced association between monoubiquitylated PCNA and REV1. In cells exposed to UV radiation, the association of REV1 with replication foci is dependent on functional UBMs. The UBMs of REV1 are shown to contribute to DNA damage tolerance and damage-induced mutagenesis in vivo.
Figures


References
-
- Baynton, K., A. Bresson-Roy, and R. P. Fuchs. 1999. Distinct roles for Rev1p and Rev7p during translesion synthesis in Saccharomyces cerevisiae. Mol. Microbiol. 34:124-133. - PubMed
-
- Bebenek, K., and T. A. Kunkel. 2004. Functions of DNA polymerases. Adv. Protein Chem. 69:137-165. - PubMed
-
- Bienko, M., C. M. Green, N. Crosetto, F. Rudolf, G. Zapart, B. Coull, P. Kannouche, G. Wider, M. Peter, A. R. Lehmann, K. Hofmann, and I. Dikic. 2005. Ubiquitin-binding domains in Y-family polymerases regulate translesion synthesis. Science 310:1821-1824. - PubMed
-
- Buerstedde, J. M., and S. Takeda. 1991. Increased ratio of targeted to random integration after transfection of chicken B cell lines. Cell 67:179-188. - PubMed
-
- Friedberg, E. C., A. R. Lehmann, and R. P. Fuchs. 2005. Trading places: how do DNA polymerases switch during translesion DNA synthesis? Mol. Cell 18:499-505. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous