

Specific isoforms of translation initiation factor 4GI show differences in translational activity
- PMID: 16982693
- PMCID: PMC1636793
- DOI: 10.1128/MCB.01248-06
Specific isoforms of translation initiation factor 4GI show differences in translational activity
Abstract
The eukaryotic initiation factor (eIF) 4GI gene locus (eIF4GI) contains three identified promoters, generating alternately spliced mRNAs, yielding a total of five eIF4GI protein isoforms. Although eIF4GI plays a critical role in mRNA recruitment to the ribosomes, little is known about the functions of the different isoforms, their partner binding capacities, or the role of the homolog, eIF4GII, in translation initiation. To directly address this, we have used short interfering RNAs (siRNAs) expressed from DNA vectors to silence the expression of eIF4GI in HeLa cells. Here we show that reduced levels of specific mRNA and eIF4GI isoforms in HeLa cells promoted aberrant morphology and a partial inhibition of translation. The latter reflected dephosphorylation of 4E-BP1 and decreased eIF4F complex levels, with no change in eIF2alpha phosphorylation. Expression of siRNA-resistant Myc-tagged eIF4GI isoforms has allowed us to show that the different isoforms exhibit significant differences in their ability to restore translation rates. Here we quantify the efficiency of eIF4GI promoter usage in mammalian cells and demonstrate that even though the longest isoform of eIF4GI (eIF4GIf) was relatively poorly expressed when reintroduced, it was more efficient at promoting the translation of cellular mRNAs than the more highly expressed shorter isoforms used in previous functional studies.
Figures
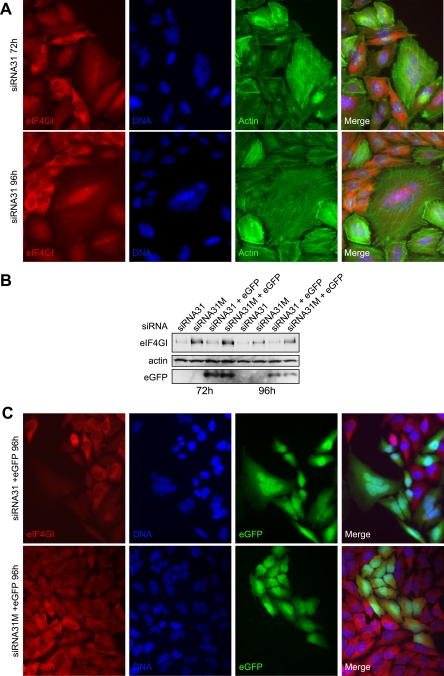

 , P < 0.001; n = 3) and comparing eIF4GIf with other isoforms of eIF4GI (, P < 0.01; ns, P = >0.05; n = 3), with error bars representing standard errors of the means.
, P < 0.001; n = 3) and comparing eIF4GIf with other isoforms of eIF4GI (, P < 0.01; ns, P = >0.05; n = 3), with error bars representing standard errors of the means.Similar articles
-
Translation of mRNAs from vesicular stomatitis virus and vaccinia virus is differentially blocked in cells with depletion of eIF4GI and/or eIF4GII.J Mol Biol. 2009 Dec 4;394(3):506-21. doi: 10.1016/j.jmb.2009.09.036. Epub 2009 Sep 19. J Mol Biol. 2009. PMID: 19769989
-
Expression of fragments of translation initiation factor eIF4GI reveals a nuclear localisation signal within the N-terminal apoptotic cleavage fragment N-FAG.J Cell Sci. 2004 May 15;117(Pt 12):2545-55. doi: 10.1242/jcs.01106. Epub 2004 May 5. J Cell Sci. 2004. PMID: 15128869
-
Cleavage of eukaryotic translation initiation factor 4GII correlates with translation inhibition during apoptosis.Cell Death Differ. 2000 Dec;7(12):1234-43. doi: 10.1038/sj.cdd.4400750. Cell Death Differ. 2000. PMID: 11175261
-
Alternative translation start sites and hidden coding potential of eukaryotic mRNAs.Bioessays. 2008 Jul;30(7):683-91. doi: 10.1002/bies.20771. Bioessays. 2008. PMID: 18536038 Review.
-
[Similar features in mechanisms of translation initiation of mRNAs in eukaryotic and prokaryotic systems].Mol Biol (Mosk). 2006 Jul-Aug;40(4):694-702. Mol Biol (Mosk). 2006. PMID: 16913228 Review. Russian.
Cited by
-
Proteomic and protein interaction network analysis of human T lymphocytes during cell-cycle entry.Mol Syst Biol. 2012 Mar 13;8:573. doi: 10.1038/msb.2012.5. Mol Syst Biol. 2012. PMID: 22415777 Free PMC article.
-
Hepatic translation control in the late-gestation fetal rat.Am J Physiol Regul Integr Comp Physiol. 2008 Aug;295(2):R558-67. doi: 10.1152/ajpregu.00091.2008. Epub 2008 Jun 18. Am J Physiol Regul Integr Comp Physiol. 2008. PMID: 18565838 Free PMC article.
-
mTOR inhibition and levels of the DNA repair protein MGMT in T98G glioblastoma cells.Mol Cancer. 2014 Jun 8;13:144. doi: 10.1186/1476-4598-13-144. Mol Cancer. 2014. PMID: 24909675 Free PMC article.
-
Reduced DEAF1 function during type 1 diabetes inhibits translation in lymph node stromal cells by suppressing Eif4g3.J Mol Cell Biol. 2013 Apr;5(2):99-110. doi: 10.1093/jmcb/mjs052. Epub 2012 Aug 24. J Mol Cell Biol. 2013. PMID: 22923498 Free PMC article.
-
Ribosome profiling reveals a functional role for autophagy in mRNA translational control.Commun Biol. 2020 Jul 17;3(1):388. doi: 10.1038/s42003-020-1090-2. Commun Biol. 2020. PMID: 32681145 Free PMC article.
References
-
- Abbott, C. M., and C. G. Proud. 2004. Translation factors: in sickness and in health. Trends Biochem. Sci. 29:25-31. - PubMed
-
- Avdulov, S., S. Li, V. Michalek, D. Burrichter, M. Peterson, D. M. Perlman, J. C. Manivel, N. Sonenberg, D. Yee, P. B. Bitterman, and V. A. Polunovsky. 2004. Activation of translation complex eIF4F is essential for the genesis and maintenance of the malignant phenotype in human mammary epithelial cells. Cancer Cell 5:553-563. - PubMed
-
- Bauer, C., I. Diesinger, N. Brass, H. Steinhart, H. Iro, and E. U. Meese. 2001. Translation initiation factor eIF-4G is immunogenic, overexpressed, and amplified in patients with squamous cell lung carcinoma. Cancer 92:822-829. - PubMed
-
- Bellsolell, L., P. F. Cho-Park, F. Poulin, N. Sonenberg, and S. K. Burley. 2006. Two structurally atypical HEAT domains in the C-terminal portion of human eIF4G support binding to eIF4A and Mnk1. Structure 14:913-923. - PubMed
-
- Belsham, G. J., and R. J. Jackson. 2000. Translation initiation on Picornavirus RNA, p. 869-900. In N. Sonenberg, J. W. B. Hershey, and M. B. Mathews (ed.), Translational control of gene expression. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases