

Interleukin (IL)-22 and IL-17 are coexpressed by Th17 cells and cooperatively enhance expression of antimicrobial peptides
- PMID: 16982811
- PMCID: PMC2118116
- DOI: 10.1084/jem.20061308
Interleukin (IL)-22 and IL-17 are coexpressed by Th17 cells and cooperatively enhance expression of antimicrobial peptides
Abstract
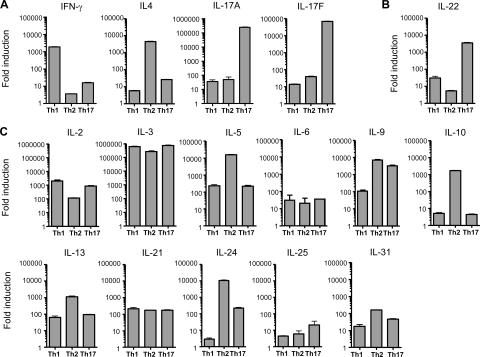
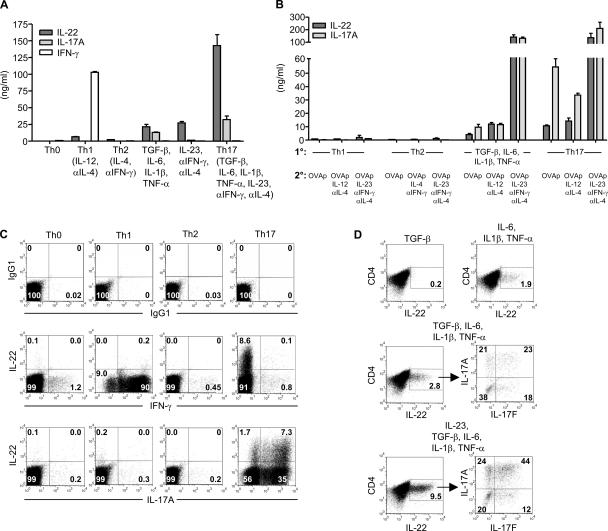
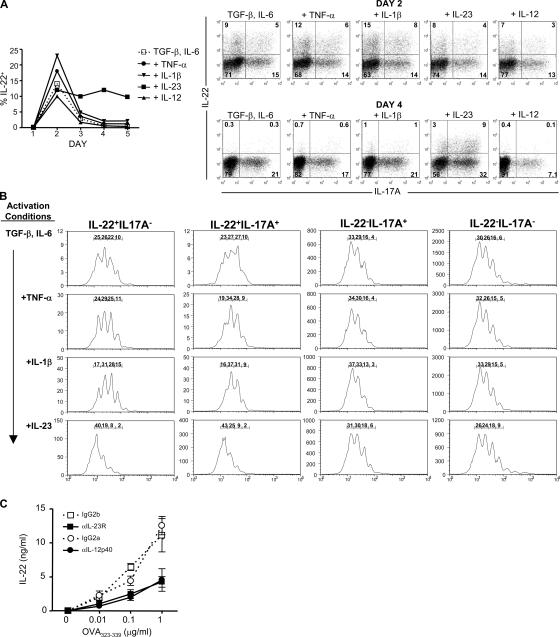
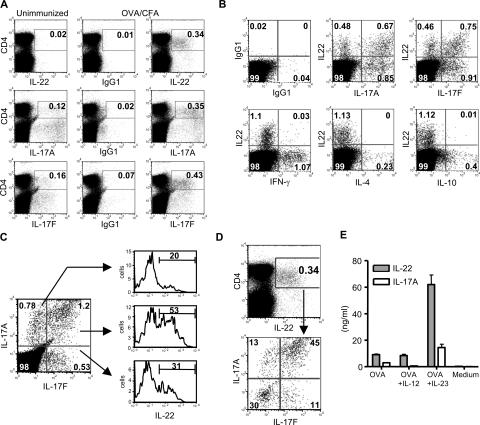
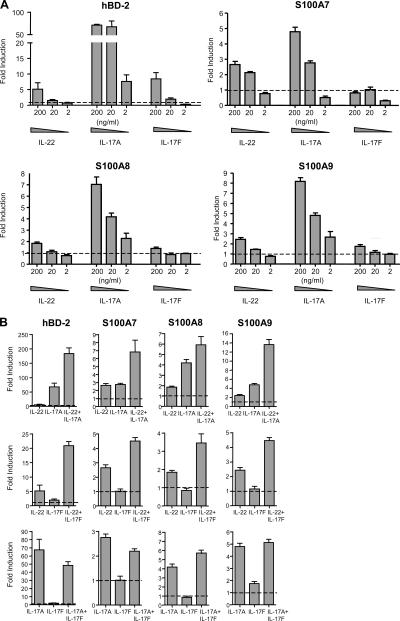
Th17 cells are a distinct lineage of effector CD4(+) T cells characterized by their production of interleukin (IL)-17. We demonstrate that Th17 cells also expressed IL-22, an IL-10 family member, at substantially higher amounts than T helper (Th)1 or Th2 cells. Similar to IL-17A, IL-22 expression was initiated by transforming growth factor beta signaling in the context of IL-6 and other proinflammatory cytokines. The subsequent expansion of IL-22-producing cells was dependent on IL-23. We further demonstrate that IL-22 was coexpressed in vitro and in vivo with both IL-17A and IL-17F. To study a functional relationship among these cytokines, we examined the expression of antimicrobial peptides by primary keratinocytes treated with combinations of IL-22, IL-17A, and IL-17F. IL-22 in conjunction with IL-17A or IL-17F synergistically induced the expression of beta-defensin 2 and S100A9 and additively enhanced the expression of S100A7 and S100A8. Collectively, we have identified IL-22 as a new cytokine expressed by Th17 cells that synergizes with IL-17A or IL-17F to regulate genes associated with skin innate immunity.
Figures
References
-
- Kolls, J.K., and A. Linden. 2004. Interleukin-17 family members and inflammation. Immunity. 21:467–476. - PubMed
-
- Aggarwal, S., N. Ghilardi, M.H. Xie, F.J. de Sauvage, and A.L. Gurney. 2003. Interleukin-23 promotes a distinct CD4 T cell activation state characterized by the production of interleukin-17. J. Biol. Chem. 278:1910–1914. - PubMed
-
- Harrington, L.E., R.D. Hatton, P.R. Mangan, H. Turner, T.L. Murphy, K.M. Murphy, and C.T. Weaver. 2005. Interleukin 17-producing CD4+ effector T cells develop via a lineage distinct from the T helper type 1 and 2 lineages. Nat. Immunol. 6:1123–1132. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
