

Crystal structure of 12-oxophytodienoate reductase 3 from tomato: self-inhibition by dimerization
- PMID: 16983071
- PMCID: PMC1586121
- DOI: 10.1073/pnas.0606603103
Crystal structure of 12-oxophytodienoate reductase 3 from tomato: self-inhibition by dimerization
Abstract
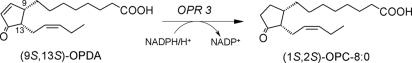
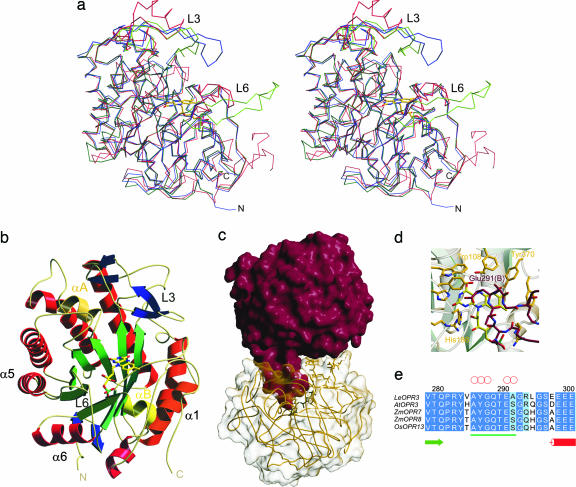
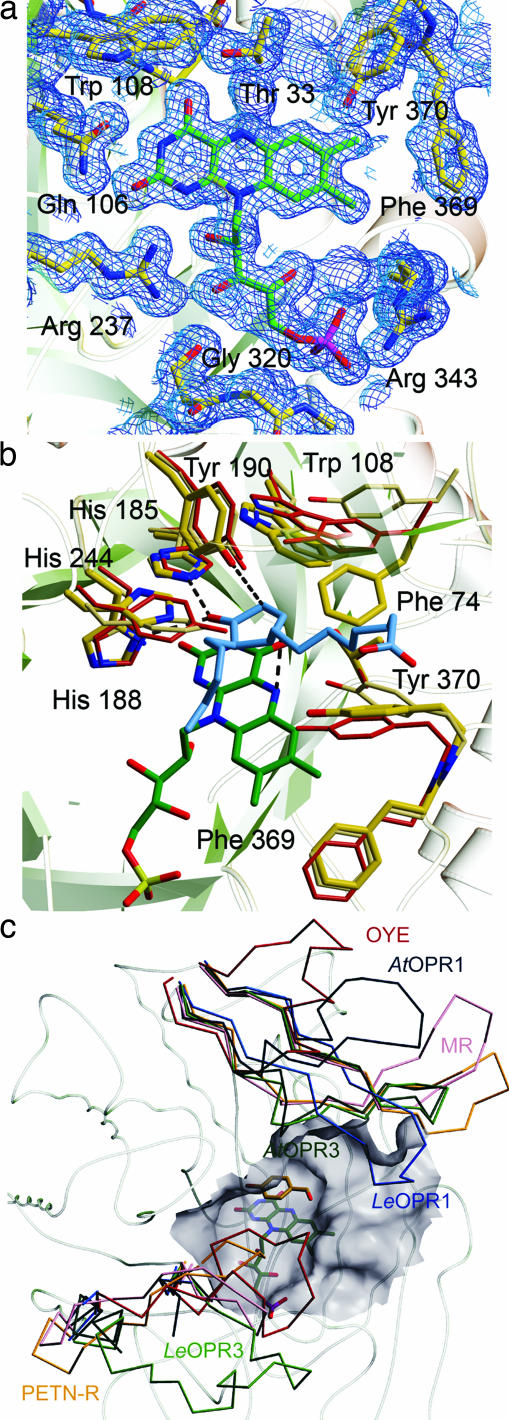
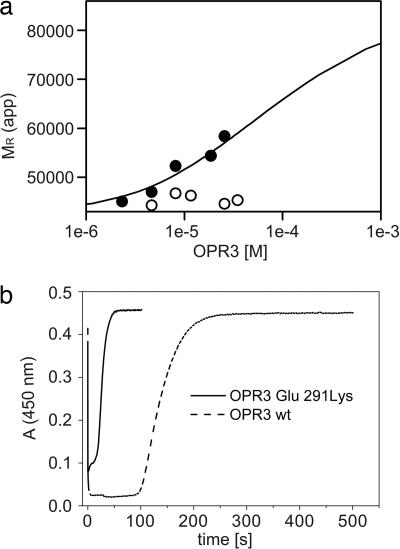
12-Oxophytodienoate reductase (OPR) 3, a homologue of old yellow enzyme (OYE), catalyzes the reduction of 9S,13S-12-oxophytodienoate to the corresponding cyclopentanone, which is subsequently converted to the plant hormone jasmonic acid (JA). JA and JA derivatives, as well as 12-oxophytodienoate and related cyclopentenones, are known to regulate gene expression in plant development and defense. Together with other oxygenated fatty acid derivatives, they form the oxylipin signature in plants, which resembles the pool of prostaglandins in animals. Here, we report the crystal structure of OPR3 from tomato and of two OPR3 mutants. Although the catalytic residues of OPR3 and related OYEs are highly conserved, several characteristic differences can be discerned in the substrate-binding regions, explaining the remarkable substrate stereoselectivity of OPR isozymes. Interestingly, OPR3 crystallized as an extraordinary self-inhibited dimer. Mutagenesis studies and biochemical analysis confirmed a weak dimerization of OPR3 in vitro, which correlated with a loss of enzymatic activity. Based on structural data of OPR3, a putative mechanism for a strong and reversible dimerization of OPR3 in vivo that involves phosphorylation of OPR3 is suggested. This mechanism could contribute to the shaping of the oxylipin signature, which is critical for fine-tuning gene expression in plants.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
