

Regulation of dendritic cell migration and adaptive immune response by leukotriene B4 receptors: a role for LTB4 in up-regulation of CCR7 expression and function
- PMID: 16985179
- PMCID: PMC1785104
- DOI: 10.1182/blood-2006-02-003665
Regulation of dendritic cell migration and adaptive immune response by leukotriene B4 receptors: a role for LTB4 in up-regulation of CCR7 expression and function
Abstract
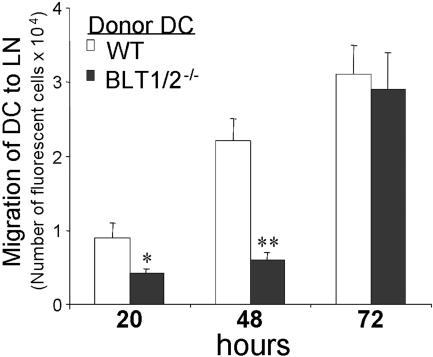
Trafficking of dendritic cells (DCs) to peripheral tissues and to secondary lymphoid organs depends on chemokines and lipid mediators. Here, we show that bone marrow-derived DCs (BM-DCs) express functional leukotriene B4 (LTB4) receptors as observed in dose-dependent chemotaxis and calcium mobilization responses. LTB4, at low concentrations, promoted the migration of immature and mature DCs to CCL19 and CCL21, which was associated with a rapid (30-minute) increase of CCR7 expression at the membrane level. At longer incubation times (6 hours), gene array analysis revealed a promoting role of LTB4, showing a significant increase of CCR7 and CCL19 mRNA levels. BM-DCs cultured from BLT1-/- or BLT1/2-/- mice showed a normal phenotype, but in vivo BLT1/2-/-DCs showed dramatic decrease in migration to the draining lymph nodes relative to wild-type (WT) DCs. Consistent with these observations, BLT1/2-/- mice showed a reduced response in a model of 2,4-dinitro-fluorobenzene (DNFB)-induced contact hypersensitivity. Adoptive transfer of 2,4-dinitrobenzene sulfonic acid (DNBS)-pulsed DCs directly implicated the defect in DC migration to lymph node with the defect in contact hypersensitivity. These results provide strong evidence for a role of LTB4 in regulating DC migration and the induction of adaptive immune responses.
Conflict of interest statement
Conflict-of-interest disclosure: The authors declare no competing financial interests.
Figures






References
-
- Banchereau J, Briere F, Caux C, et al. Immunobiology of dendritic cells. Annu Rev Immunol. 2000;18:767–811. - PubMed
-
- Steinman RM, Hawiger D, Nussenzweig MC. Tolerogenic dendritic cells. Annu Rev Immunol. 2003;21:685–711. - PubMed
-
- Sozzani S, Allavena P, D'Amico G, et al. Differential regulation of chemokine receptors during dendritic cell maturation: a model for their trafficking properties. J Immunol. 1998;161:1083–1086. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
