

Yeast sphingolipids do not need to contain very long chain fatty acids
- PMID: 16987101
- PMCID: PMC1698682
- DOI: 10.1042/BJ20061128
Yeast sphingolipids do not need to contain very long chain fatty acids
Abstract
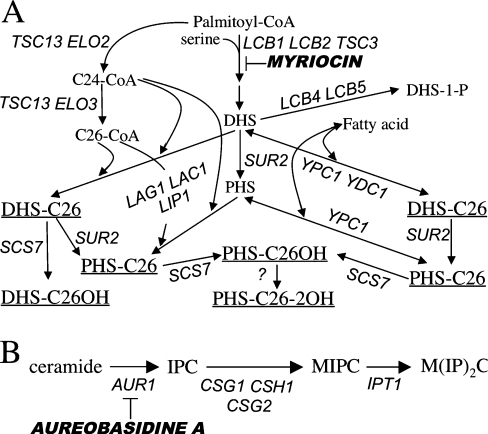
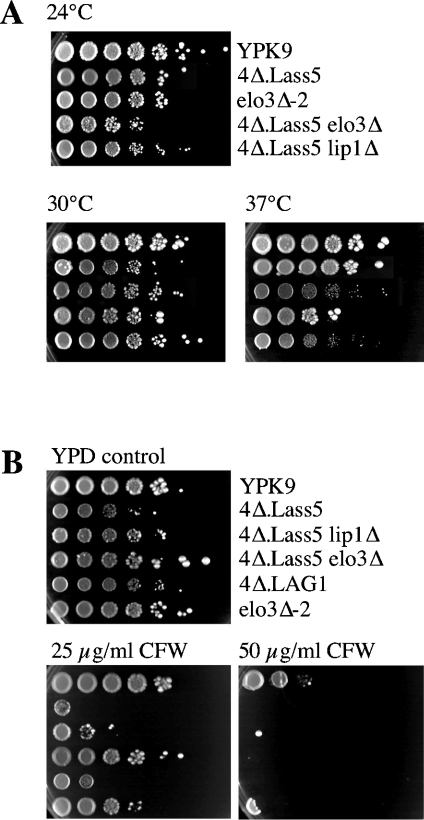
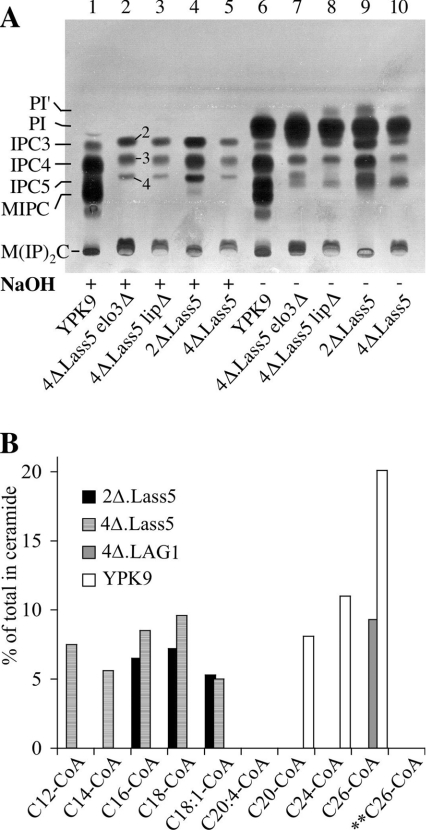
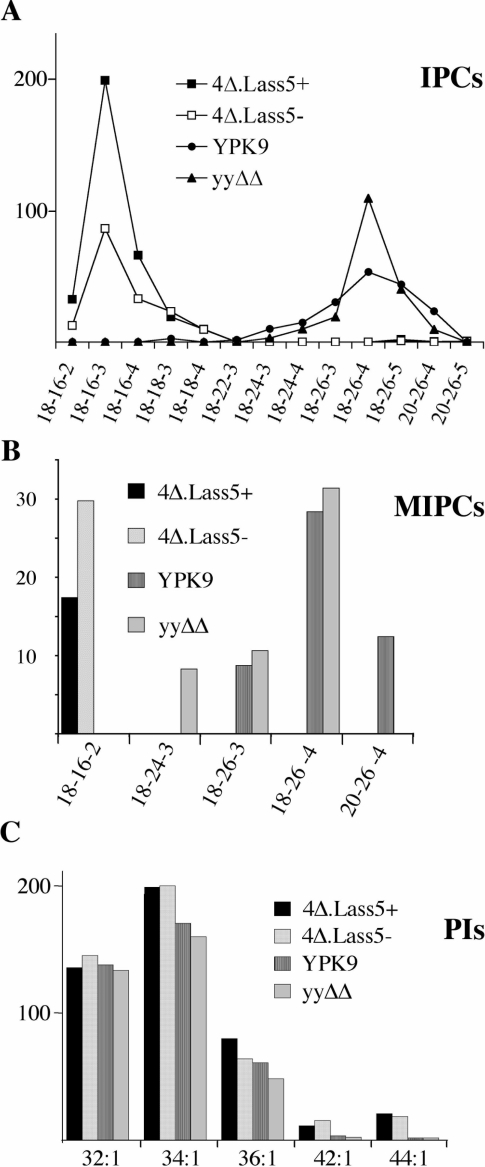
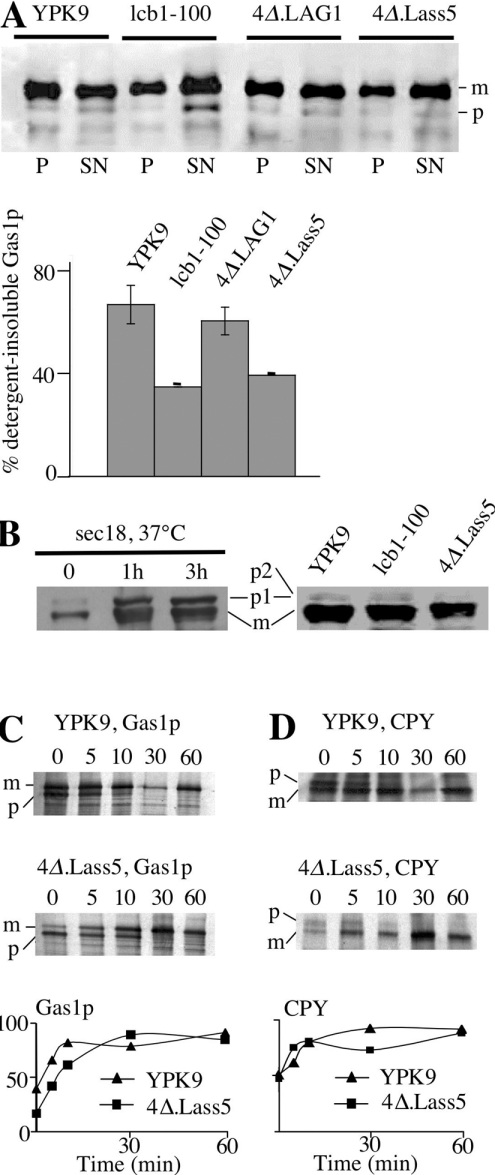
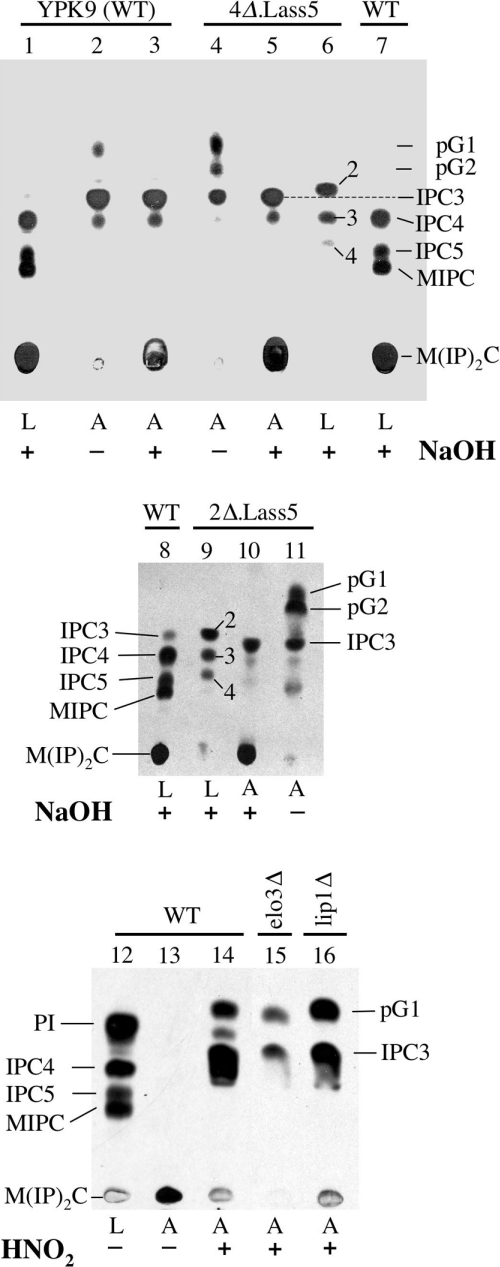
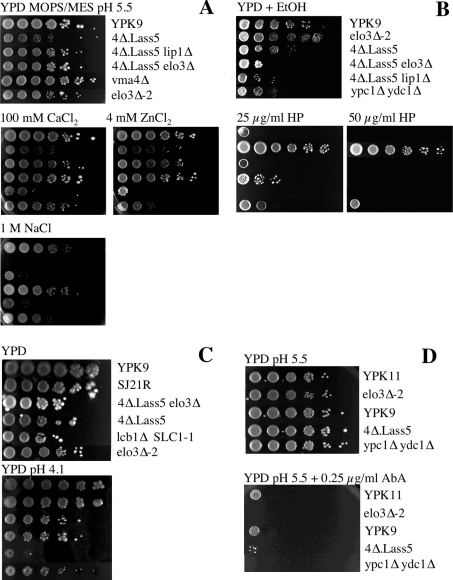
Synthesis of VLCFAs (very long chain fatty acids) and biosynthesis of DHS (dihydrosphingosine) both are of vital importance for Saccharomyces cerevisiae. The bulk of VLCFAs and DHS are used for ceramide synthesis by the Lag1p (longevity-assurance gene 1)/Lac1p (longevity-assurance gene cognate 1)/Lip1p (Lag1p/Lac1p interacting protein) ceramide synthase. LAG1 and LAC1 are redundant but LIP1 is essential. Here we show that 4Delta (lag1Deltalac1Deltaypc1Deltaydc1Delta) cells devoid of all known endogenous ceramide synthesis pathways are unviable but can be rescued by the expression of Lass5, a mouse LAG1 homologue. Ceramide synthase activity of 4Delta.Lass5 cells only utilizes C16 and C18 fatty acids and does not require the help of Lip1p, an essential cofactor of Lag1p/Lac1p. HPLC-electrospray ionization-MS/MS analysis demonstrated that in IPCs (inositolphosphorylceramides) of 4Delta.Lass5, the very long chain fatty acids (C26 and C24) account for <1% instead of the normal >97%. Notwithstanding, IPCs incorporated into glycosylphosphatidylinositol anchors of 4Delta.Lass5 show normal mobility on TLC and the ceramide- and raft-dependent traffic of Gas1p (glycophospholipid-anchored surface protein) from endoplasmic reticulum to Golgi remains almost normal. Moreover, the biosynthesis of C24:0 fatty acids remains essential. Thus, C(24:0) and dihydrosphingosine are both necessary for survival of yeast cells even if they utilize C16 and C18 fatty acids for sphingolipid biosynthesis.
Figures
References
-
- Dickson R. C., Sumanasekera C., Lester R. L. Functions and metabolism of sphingolipids in Saccharomyces cerevisiae. Prog. Lipid Res. 2006;45:447–465. - PubMed
-
- Smith S. W., Lester R. L. Inositol phosphorylceramide, a novel substance and the chief member of a major group of yeast sphingolipids containing a single inositol phosphate. J. Biol. Chem. 1974;249:3395–3405. - PubMed
-
- Oh C. S., Toke D. A., Mandala S., Martin C. E. ELO2 and ELO3, homologues of the Saccharomyces cerevisiae ELO1 gene, function in fatty acid elongation and are required for sphingolipid formation. J. Biol. Chem. 1997;272:17376–17384. - PubMed
-
- Sims K. J., Spassieva S. D., Voit E. O., Obeid L. M. Yeast sphingolipid metabolism: clues and connections. Biochem. Cell Biol. 2004;82:45–61. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases