

Rho GTPase activity zones and transient contractile arrays
- PMID: 16998826
- PMCID: PMC4364130
- DOI: 10.1002/bies.20477
Rho GTPase activity zones and transient contractile arrays
Abstract
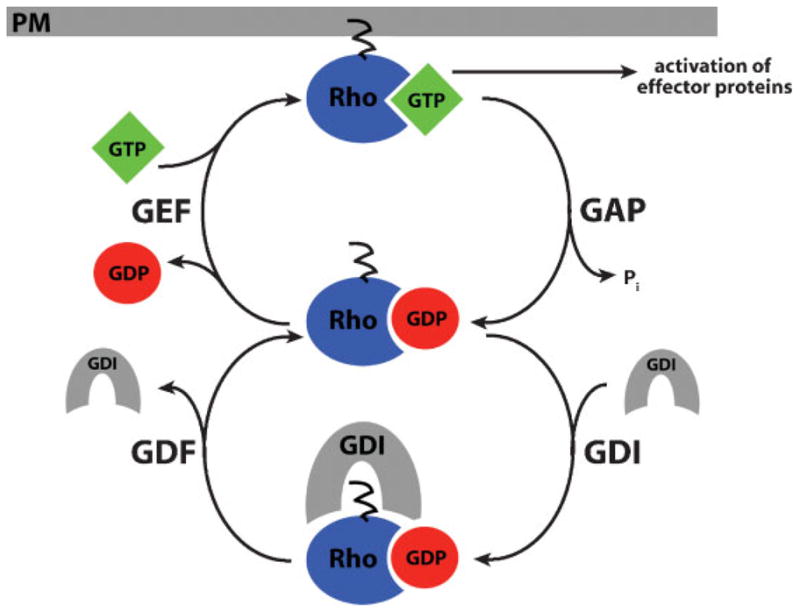
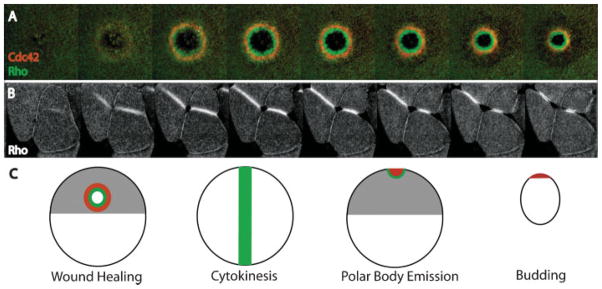
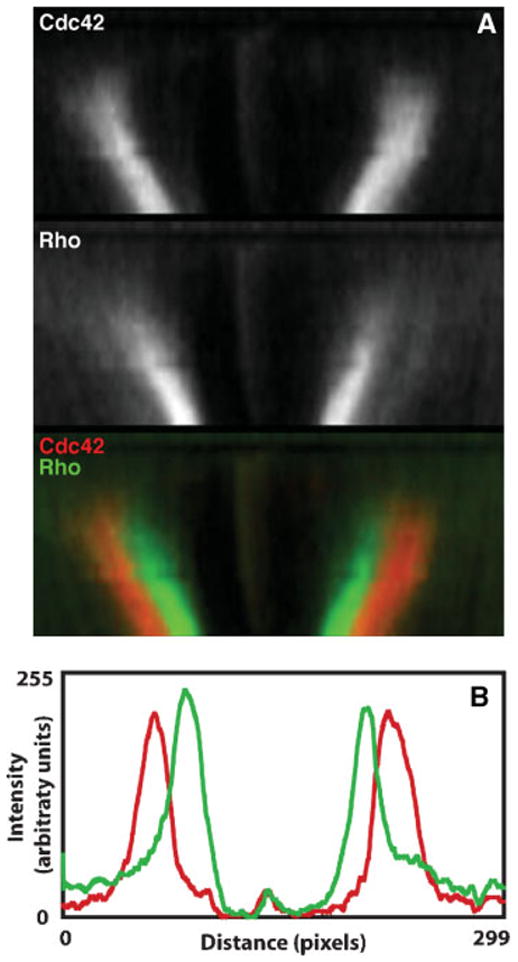
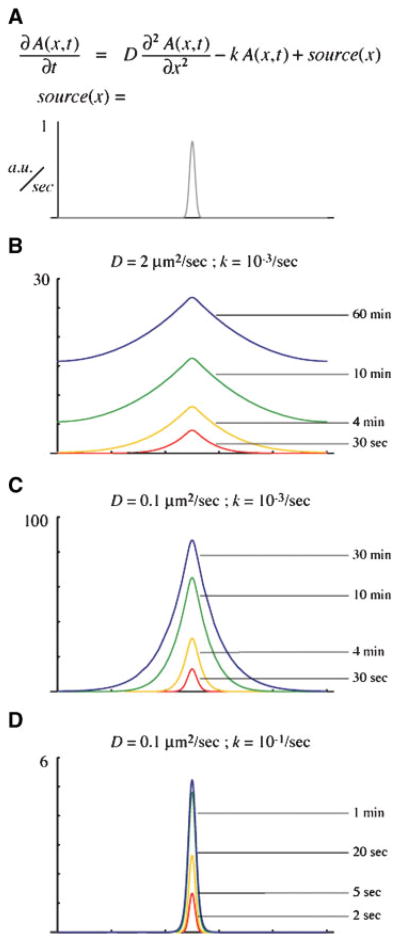
The Rho GTPases-Rho, Rac and Cdc42-act as molecular switches, cycling between an active GTP-bound state and an inactive GDP-bound state, to regulate the actin cytoskeleton. It has recently become apparent that the Rho GTPases can be activated in subcellular zones that appear semi-stable, yet are dynamically maintained. These Rho GTPase activity zones are associated with a variety of fundamental biological processes including symmetric and asymmetric cytokinesis and cellular wound repair. Here we review the basic features of Rho GTPase activity zones, suggest that these zones represent a fundamental signaling mechanism, and discuss the implications of zone properties from the perspective of both their function and how they are likely to be controlled.
(c) 2006 Wiley Periodicals, Inc.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
