

Dimerization of the I kappa B kinase-binding domain of NEMO is required for tumor necrosis factor alpha-induced NF-kappa B activity
- PMID: 17000764
- PMCID: PMC1698548
- DOI: 10.1128/MCB.00478-06
Dimerization of the I kappa B kinase-binding domain of NEMO is required for tumor necrosis factor alpha-induced NF-kappa B activity
Abstract
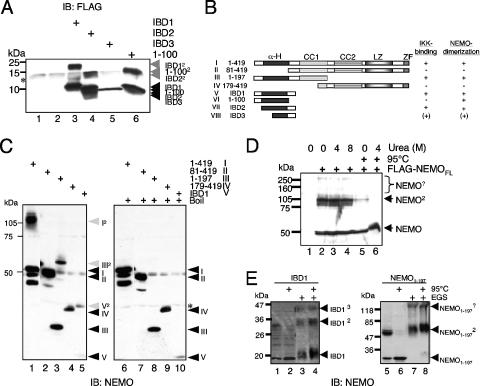
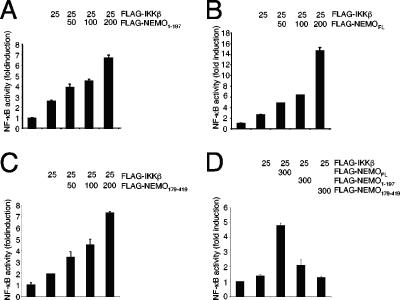
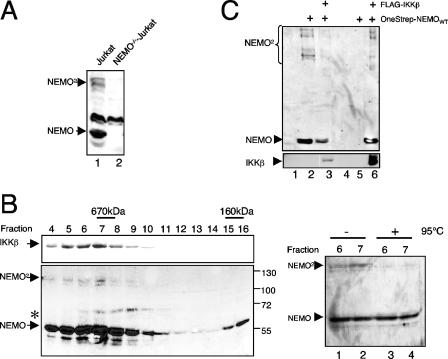
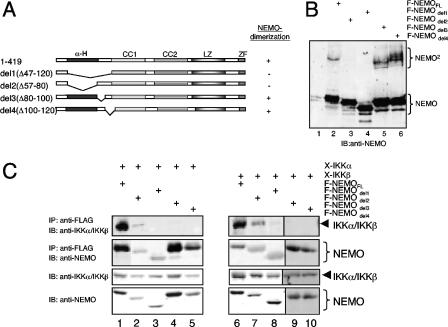
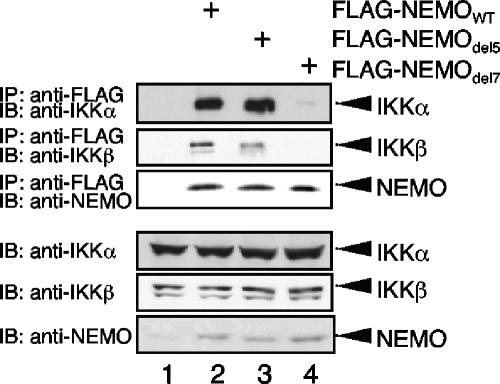
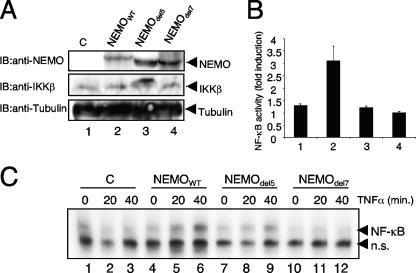
Previous studies have demonstrated that peptides corresponding to a six-amino-acid NEMO-binding domain from the C terminus of IkappaB kinase alpha (IKKalpha) and IKKbeta can disrupt the IKK complex and block NF-kappaB activation. We have now mapped and characterized the corresponding amino-terminal IKK-binding domain (IBD) of NEMO. Peptides corresponding to the IBD were efficiently recruited to the IKK complex but displayed only a weak inhibitory potential on cytokine-induced NF-kappaB activity. This is most likely due to the formation of sodium dodecyl sulfate- and urea-resistant NEMO dimers through a dimerization domain at the amino terminus of NEMO that overlaps with the region responsible for binding to IKKs. Mutational analysis revealed different alpha-helical subdomains within an amino-terminal coiled-coil region are important for NEMO dimerization and IKKbeta binding. Furthermore, NEMO dimerization is required for the tumor necrosis factor alpha-induced NF-kappaB activation, even when interaction with the IKKs is unaffected. Hence, our data provide novel insights into the role of the amino terminus of NEMO for the architecture of the IKK complex and its activation.
Figures



Similar articles
-
Phosphorylation of serine 68 in the IkappaB kinase (IKK)-binding domain of NEMO interferes with the structure of the IKK complex and tumor necrosis factor-alpha-induced NF-kappaB activity.J Biol Chem. 2008 Jan 4;283(1):76-86. doi: 10.1074/jbc.M708856200. Epub 2007 Oct 31. J Biol Chem. 2008. PMID: 17977820
-
NEMO-binding domains of both IKKalpha and IKKbeta regulate IkappaB kinase complex assembly and classical NF-kappaB activation.J Biol Chem. 2009 Oct 2;284(40):27596-608. doi: 10.1074/jbc.M109.047563. Epub 2009 Aug 7. J Biol Chem. 2009. PMID: 19666475 Free PMC article.
-
Dimer of arfaptin 2 regulates NF-κB signaling by interacting with IKKβ/NEMO and inhibiting IKKβ kinase activity.Cell Signal. 2015 Nov;27(11):2173-81. doi: 10.1016/j.cellsig.2015.08.012. Epub 2015 Aug 18. Cell Signal. 2015. PMID: 26296658
-
Regulation and function of IKK and IKK-related kinases.Sci STKE. 2006 Oct 17;2006(357):re13. doi: 10.1126/stke.3572006re13. Sci STKE. 2006. PMID: 17047224 Review.
-
Protein-protein interactions involving IKKgamma (NEMO) that promote the activation of NF-kappaB.J Cell Physiol. 2010 Jun;223(3):558-61. doi: 10.1002/jcp.22105. J Cell Physiol. 2010. PMID: 20301198 Review.
Cited by
-
NF-κB, the first quarter-century: remarkable progress and outstanding questions.Genes Dev. 2012 Feb 1;26(3):203-34. doi: 10.1101/gad.183434.111. Genes Dev. 2012. PMID: 22302935 Free PMC article. Review.
-
The Trypanosoma cruzi protease cruzain mediates immune evasion.PLoS Pathog. 2011 Sep;7(9):e1002139. doi: 10.1371/journal.ppat.1002139. Epub 2011 Sep 1. PLoS Pathog. 2011. PMID: 21909255 Free PMC article.
-
NEMO oligomerization and its ubiquitin-binding properties.Biochem J. 2009 Jun 26;421(2):243-51. doi: 10.1042/BJ20090427. Biochem J. 2009. PMID: 19422324 Free PMC article.
-
Inhibition of NF-kappaB activation with designed ankyrin-repeat proteins targeting the ubiquitin-binding/oligomerization domain of NEMO.Protein Sci. 2007 Sep;16(9):2013-22. doi: 10.1110/ps.072924907. Protein Sci. 2007. PMID: 17766391 Free PMC article.
-
Intermolecular disulfide bond formation in the NEMO dimer requires Cys54 and Cys347.Biochem Biophys Res Commun. 2008 Feb 29;367(1):103-8. doi: 10.1016/j.bbrc.2007.12.123. Epub 2007 Dec 28. Biochem Biophys Res Commun. 2008. PMID: 18164680 Free PMC article.
References
-
- Agou, F., G. Courtois, J. Chiaravalli, F. Baleux, Y. M. Coic, F. Traincard, A. Israel, and M. Veron. 2004. Inhibition of NF-kappa B activation by peptides targeting NEMO oligomerization. J. Biol. Chem. 279:54248-54257. - PubMed
-
- Agou, F., F. Traincard, E. Vinolo, G. Courtois, S. Yamaoka, A. Israel, and M. Veron. 2004. The trimerization domain of NEMO is composed of the interacting C-terminal CC2 and LZ coiled-coil subdomains. J. Biol. Chem. 279:27861-27869. - PubMed
-
- Agou, F., F. Ye, S. Goffinont, G. Courtois, S. Yamaoka, A. Israel, and M. Veron. 2002. NEMO trimerizes through its coiled-coil C-terminal domain. J. Biol. Chem. 277:17464-17475. - PubMed
-
- Brummelkamp, T. R., S. M. Nijman, A. M. Dirac, and R. Bernards. 2003. Loss of the cylindromatosis tumour suppressor inhibits apoptosis by activating NF-kappaB. Nature 424:797-801. - PubMed
-
- Carter, R. S., K. N. Pennington, B. J. Ungurait, and D. W. Ballard. 2003. In vivo identification of inducible phosphoacceptors in the IKKgamma/NEMO subunit of human IkappaB kinase. J. Biol. Chem. 278:19642-19648. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous