

The Ral/exocyst effector complex counters c-Jun N-terminal kinase-dependent apoptosis in Drosophila melanogaster
- PMID: 17000765
- PMCID: PMC1636832
- DOI: 10.1128/MCB.00506-06
The Ral/exocyst effector complex counters c-Jun N-terminal kinase-dependent apoptosis in Drosophila melanogaster
Abstract
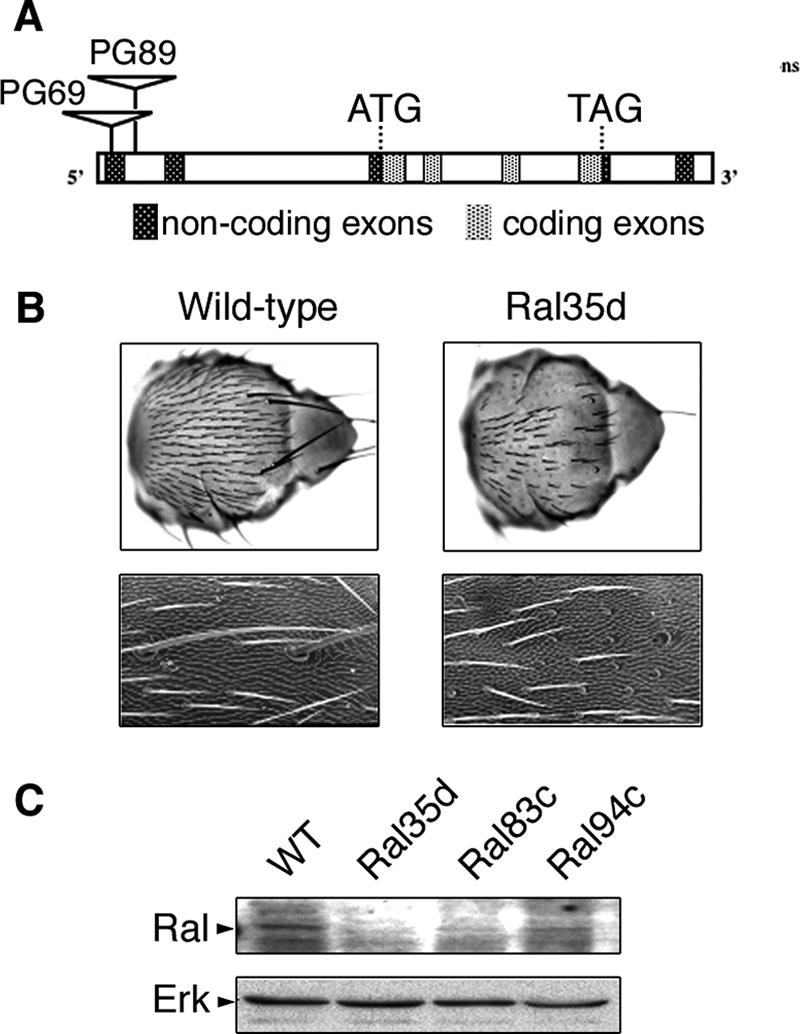
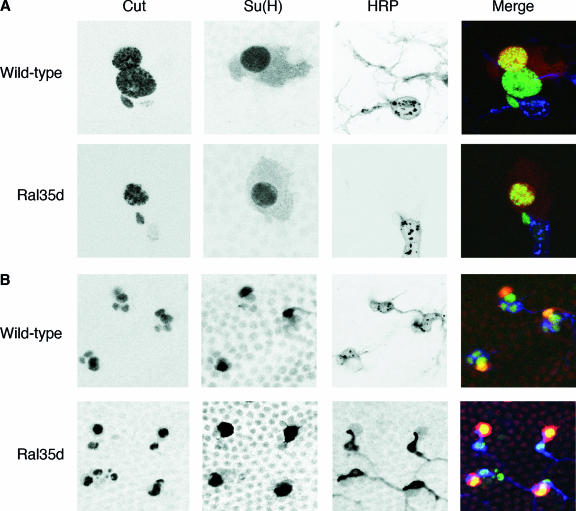
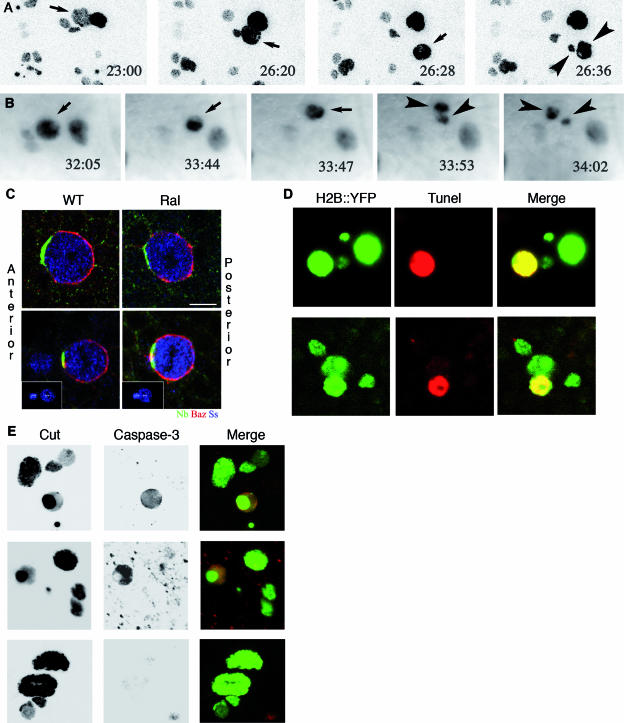
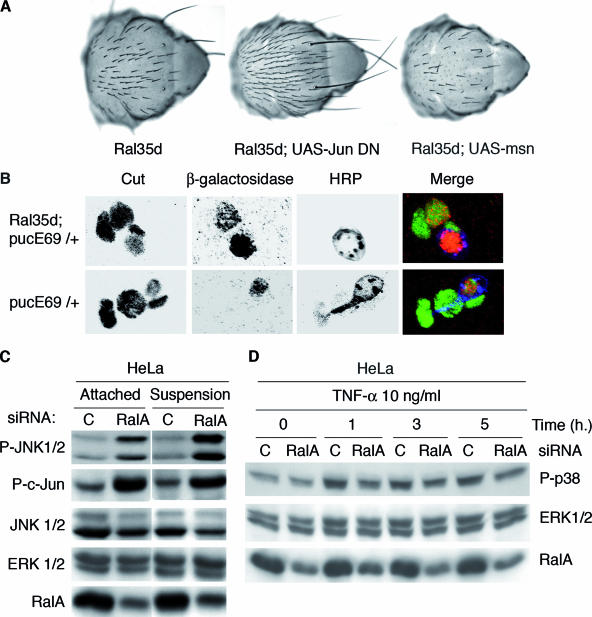
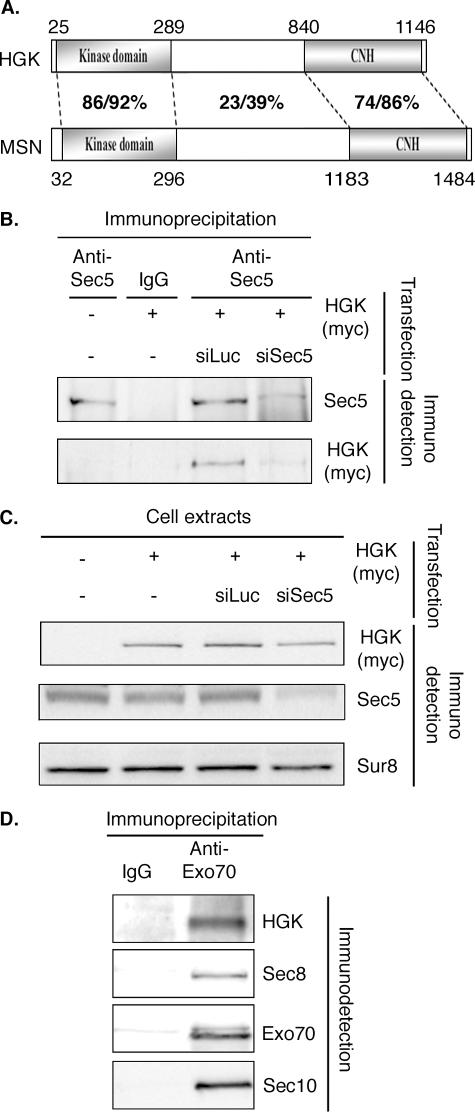
Ral GTPase activity is a crucial cell-autonomous factor supporting tumor initiation and progression. To decipher pathways impacted by Ral, we have generated null and hypomorph alleles of the Drosophila melanogaster Ral gene. Ral null animals were not viable. Reduced Ral expression in cells of the sensory organ lineage had no effect on cell division but led to postmitotic cell-specific apoptosis. Genetic epistasis and immunofluorescence in differentiating sensory organs suggested that Ral activity suppresses c-Jun N-terminal kinase (JNK) activation and induces p38 mitogen-activated protein (MAP) kinase activation. HPK1/GCK-like kinase (HGK), a MAP kinase kinase kinase kinase that can drive JNK activation, was found as an exocyst-associated protein in vivo. The exocyst is a Ral effector, and the epistasis between mutants of Ral and of msn, the fly ortholog of HGK, suggest the functional relevance of an exocyst/HGK interaction. Genetic analysis also showed that the exocyst is required for the execution of Ral function in apoptosis. We conclude that in Drosophila Ral counters apoptotic programs to support cell fate determination by acting as a negative regulator of JNK activity and a positive activator of p38 MAP kinase. We propose that the exocyst complex is Ral executioner in the JNK pathway and that a cascade from Ral to the exocyst to HGK would be a molecular basis of Ral action on JNK.
Figures
Similar articles
-
The Drosophila Ral GTPase regulates developmental cell shape changes through the Jun NH(2)-terminal kinase pathway.J Cell Biol. 1999 Jul 26;146(2):361-72. doi: 10.1083/jcb.146.2.361. J Cell Biol. 1999. PMID: 10427090 Free PMC article.
-
A novel human STE20-related protein kinase, HGK, that specifically activates the c-Jun N-terminal kinase signaling pathway.J Biol Chem. 1999 Jan 22;274(4):2118-25. doi: 10.1074/jbc.274.4.2118. J Biol Chem. 1999. PMID: 9890973
-
[The role of mitogen-activated protein kinase cascades in inhibition of proliferation in human prostate carcinoma cells by raloxifene: an in vitro experiment].Zhonghua Yi Xue Za Zhi. 2008 Jan 22;88(4):271-5. Zhonghua Yi Xue Za Zhi. 2008. PMID: 18361842 Chinese.
-
A signalling cascade for Ral.Small GTPases. 2022 Jan;13(1):128-135. doi: 10.1080/21541248.2021.1917953. Epub 2021 May 6. Small GTPases. 2022. PMID: 33956571 Free PMC article. Review.
-
Stress signaling in Drosophila.Oncogene. 1999 Nov 1;18(45):6172-82. doi: 10.1038/sj.onc.1203125. Oncogene. 1999. PMID: 10557109 Review.
Cited by
-
Exocyst dynamics during vesicle tethering and fusion.Nat Commun. 2018 Dec 3;9(1):5140. doi: 10.1038/s41467-018-07467-5. Nat Commun. 2018. PMID: 30510181 Free PMC article.
-
Exocyst Complex Member EXOC5 Is Required for Survival of Hair Cells and Spiral Ganglion Neurons and Maintenance of Hearing.Mol Neurobiol. 2018 Aug;55(8):6518-6532. doi: 10.1007/s12035-017-0857-z. Epub 2018 Jan 11. Mol Neurobiol. 2018. PMID: 29327200 Free PMC article.
-
The small GTPase RALA controls c-Jun N-terminal kinase-mediated FOXO activation by regulation of a JIP1 scaffold complex.J Biol Chem. 2013 Jul 26;288(30):21729-41. doi: 10.1074/jbc.M113.463885. Epub 2013 Jun 14. J Biol Chem. 2013. PMID: 23770673 Free PMC article.
-
RalA and RalB proteins are ubiquitinated GTPases, and ubiquitinated RalA increases lipid raft exposure at the plasma membrane.J Biol Chem. 2012 Aug 24;287(35):29397-405. doi: 10.1074/jbc.M112.357764. Epub 2012 Jun 14. J Biol Chem. 2012. PMID: 22700969 Free PMC article.
-
Distinct roles of RalA and RalB in the progression of cytokinesis are supported by distinct RalGEFs.EMBO J. 2008 Sep 17;27(18):2375-87. doi: 10.1038/emboj.2008.166. Epub 2008 Aug 28. EMBO J. 2008. PMID: 18756269 Free PMC article.
References
-
- Adachi-Yamada, T., M. Nakamura, K. Irie, Y. Tomoyasu, Y. Sano, E. Mori, S. Goto, N. Ueno, Y. Nishida, and K. Matsumoto. 1999. p38 mitogen-activated protein kinase can be involved in transforming growth factor β superfamily signal transduction in Drosophila wing morphogenesis. Mol. Cell. Biol. 19:2322-2329. - PMC - PubMed
-
- Agapova, L. S., J. L. Volodina, P. M. Chumakov, and B. P. Kopnin. 2004. Activation of Ras-Ral pathway attenuates p53-independent DNA damage G2 checkpoint. J. Biol. Chem. 279:36382-36389. - PubMed
-
- Bellaiche, Y., M. Gho, J. A. Kaltschmidt, A. H. Brand, and F. Schweisguth. 2001. Frizzled regulates localization of cell-fate determinants and mitotic spindle rotation during asymmetric cell division. Nat. Cell Biol. 3:50-57. - PubMed
-
- Bellaiche, Y., A. Radovic, D. F. Woods, C. D. Hough, M. L. Parmentier, C. J. O'Kane, P. J. Bryant, and F. Schweisguth. 2001. The partner of inscuteable/discs-large complex is required to establish planar polarity during asymmetric cell division in Drosophila. Cell 106:355-366. - PubMed
-
- Blochlinger, K., R. Bodmer, L. Y. Jan, and Y. N. Jan. 1990. Patterns of expression of cut, a protein required for external sensory organ development in wild-type and cut mutant Drosophila embryos. Genes Dev. 4:1322-1331. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous