

Flow-enhanced adhesion regulated by a selectin interdomain hinge
- PMID: 17000883
- PMCID: PMC2064400
- DOI: 10.1083/jcb.200606056
Flow-enhanced adhesion regulated by a selectin interdomain hinge
Abstract
L-selectin requires a threshold shear to enable leukocytes to tether to and roll on vascular surfaces. Transport mechanisms govern flow-enhanced tethering, whereas force governs flow-enhanced rolling by prolonging the lifetimes of L-selectin-ligand complexes (catch bonds). Using selectin crystal structures, molecular dynamics simulations, site-directed mutagenesis, single-molecule force and kinetics experiments, Monte Carlo modeling, and flow chamber adhesion studies, we show that eliminating a hydrogen bond to increase the flexibility of an interdomain hinge in L-selectin reduced the shear threshold for adhesion via two mechanisms. One affects the on-rate by increasing tethering through greater rotational diffusion. The other affects the off-rate by strengthening rolling through augmented catch bonds with longer lifetimes at smaller forces. By forcing open the hinge angle, ligand may slide across its interface with L-selectin to promote rebinding, thereby providing a mechanism for catch bonds. Thus, allosteric changes remote from the ligand-binding interface regulate both bond formation and dissociation.
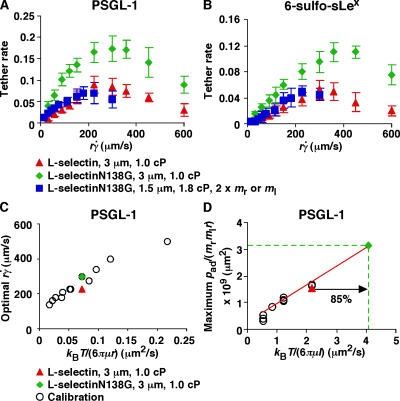
Figures






Comment in
-
For catch bonds, it all hinges on the interdomain region.J Cell Biol. 2006 Sep 25;174(7):911-3. doi: 10.1083/jcb.200609029. J Cell Biol. 2006. PMID: 17000873 Free PMC article. Review.
References
-
- Ajzenberg, N., A.S. Ribba, G. Rastegar-Lari, D. Meyer, and D. Baruch. 2000. Effect of recombinant von Willebrand factor reproducing type 2B or type 2M mutations on shear-induced platelet aggregation. Blood. 95:3796–3803. - PubMed
-
- Bell, G.I. 1978. Models for the specific adhesion of cells to cells: a theoretical framework for adhesion mediated by reversible bonds between cell surface molecules. Science. 200:618–627. - PubMed
-
- Doggett, T.A., G. Girdhar, A. Lawshe, J.L. Miller, I.J. Laurenzi, S.L. Diamond, and T.G. Diacovo. 2003. Alterations in the intrinsic properties of the GPIbα-VWF tether bond define the kinetics of the platelet-type von Willebrand disease mutation, Gly233Val. Blood. 102:152–160. - PubMed
-
- Dwir, O., G.S. Kansas, and R. Alon. 2000. An activated L-selectin mutant with conserved equilibrium binding properties but enhanced ligand recognition under shear flow. J. Biol. Chem. 275:18682–18691. - PubMed
