

Intra- and intersubunit cooperativity in activation of BK channels by Ca2+
- PMID: 17001085
- PMCID: PMC2151572
- DOI: 10.1085/jgp.200609486
Intra- and intersubunit cooperativity in activation of BK channels by Ca2+
Abstract
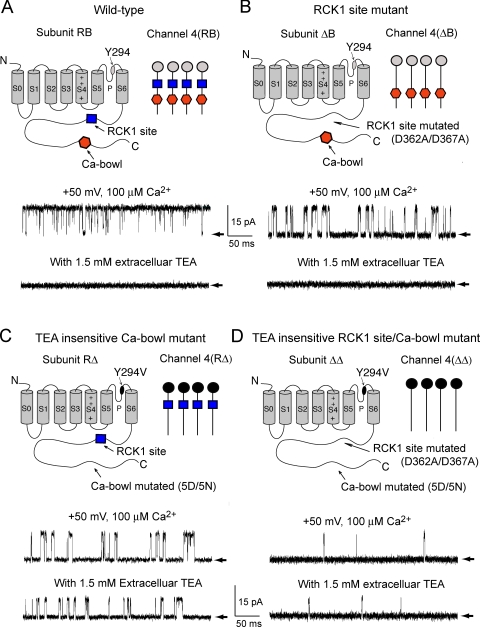
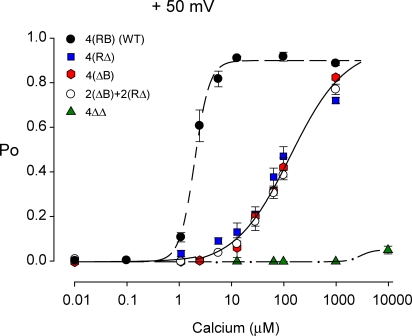
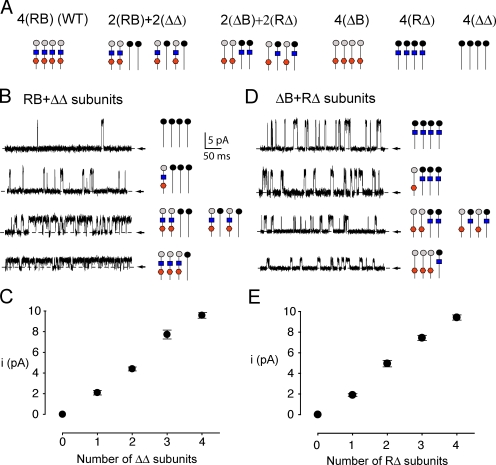
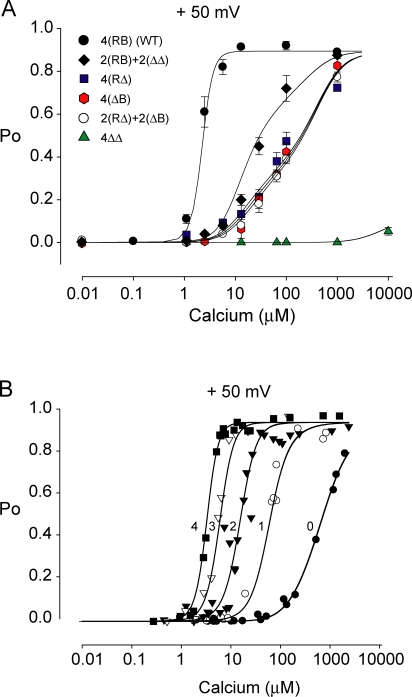
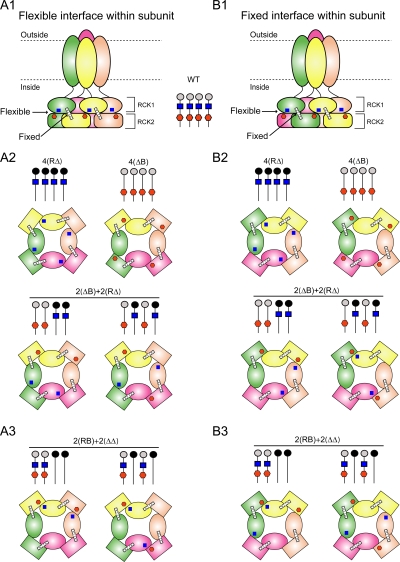
The activation of BK channels by Ca(2+) is highly cooperative, with small changes in intracellular Ca(2+) concentration having large effects on open probability (Po). Here we examine the mechanism of cooperative activation of BK channels by Ca(2+). Each of the four subunits of BK channels has a large intracellular COOH terminus with two different high-affinity Ca(2+) sensors: an RCK1 sensor (D362/D367) located on the RCK1 (regulator of conductance of K(+)) domain and a Ca-bowl sensor located on or after the RCK2 domain. To determine interactions among these Ca(2+) sensors, we examine channels with eight different configurations of functional high-affinity Ca(2+) sensors on the four subunits. We find that the RCK1 sensor and Ca bowl contribute about equally to Ca(2+) activation of the channel when there is only one high-affinity Ca(2+) sensor per subunit. We also find that an RCK1 sensor and a Ca bowl on the same subunit are much more effective in increasing Po than when they are on different subunits, indicating positive intrasubunit cooperativity. If it is assumed that BK channels have a gating ring similar to MthK channels with alternating RCK1 and RCK2 domains and that the Ca(2+) sensors act at the flexible (rather than fixed) interfaces between RCK domains, then a comparison of the distribution of Ca(2+) sensors with the observed responses suggest that the interface between RCK1 and RCK2 domains on the same subunit is flexible. On this basis, intrasubunit cooperativity arises because two high-affinity Ca(2+) sensors acting across a flexible interface are more effective in opening the channel than when acting at separate interfaces. An allosteric model incorporating intrasubunit cooperativity nested within intersubunit cooperativity could approximate the Po vs. Ca(2+) response for eight possible subunit configurations of the high-affinity Ca(2+) sensors as well as for three additional configurations from a previous study.
Figures

Comment in
-
A tail of multiple calcium-sensing domains.J Gen Physiol. 2006 Oct;128(4):387-8. doi: 10.1085/jgp.200609536. J Gen Physiol. 2006. PMID: 17001084 Free PMC article. No abstract available.
References
-
- Adelman, J.P., K.Z. Shen, M.P. Kavanaugh, R.A. Warren, Y.N. Wu, A. Lagrutta, C.T. Bond, and R.A. North. 1992. Calcium-activated potassium channels expressed from cloned complementary DNAs. Neuron. 9:209–216. - PubMed
-
- Atkinson, N.S., G.A. Robertson, and B. Ganetzky. 1991. A component of calcium-activated potassium channels encoded by the Drosophila slo locus. Science. 253:551–555. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
