

Redesign of Schistosoma mansoni NAD+ catabolizing enzyme: active site H103W mutation restores ADP-ribosyl cyclase activity
- PMID: 17002287
- PMCID: PMC2546491
- DOI: 10.1021/bi060930g
Redesign of Schistosoma mansoni NAD+ catabolizing enzyme: active site H103W mutation restores ADP-ribosyl cyclase activity
Abstract
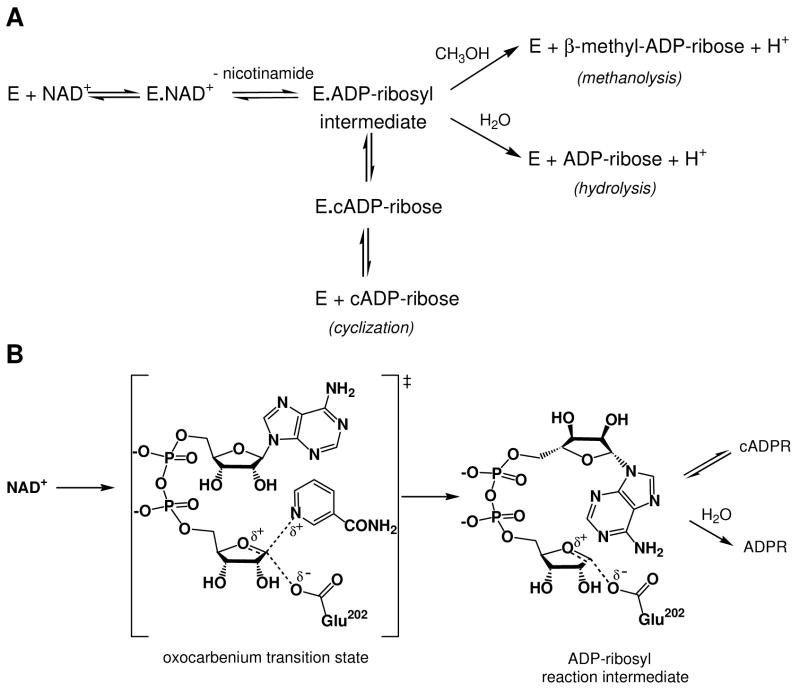
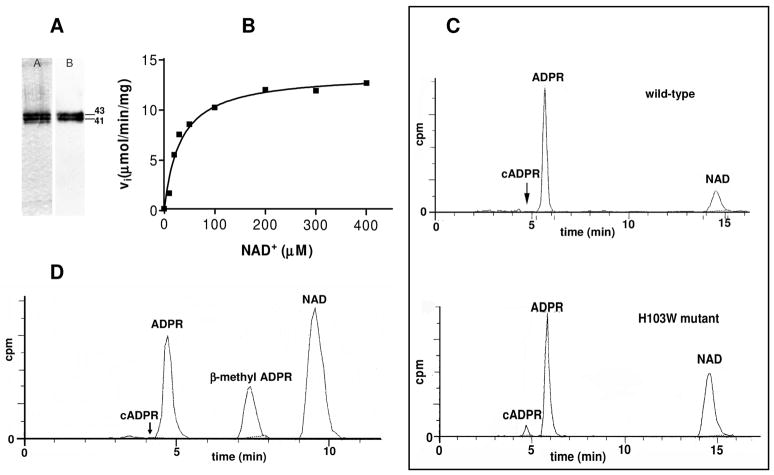
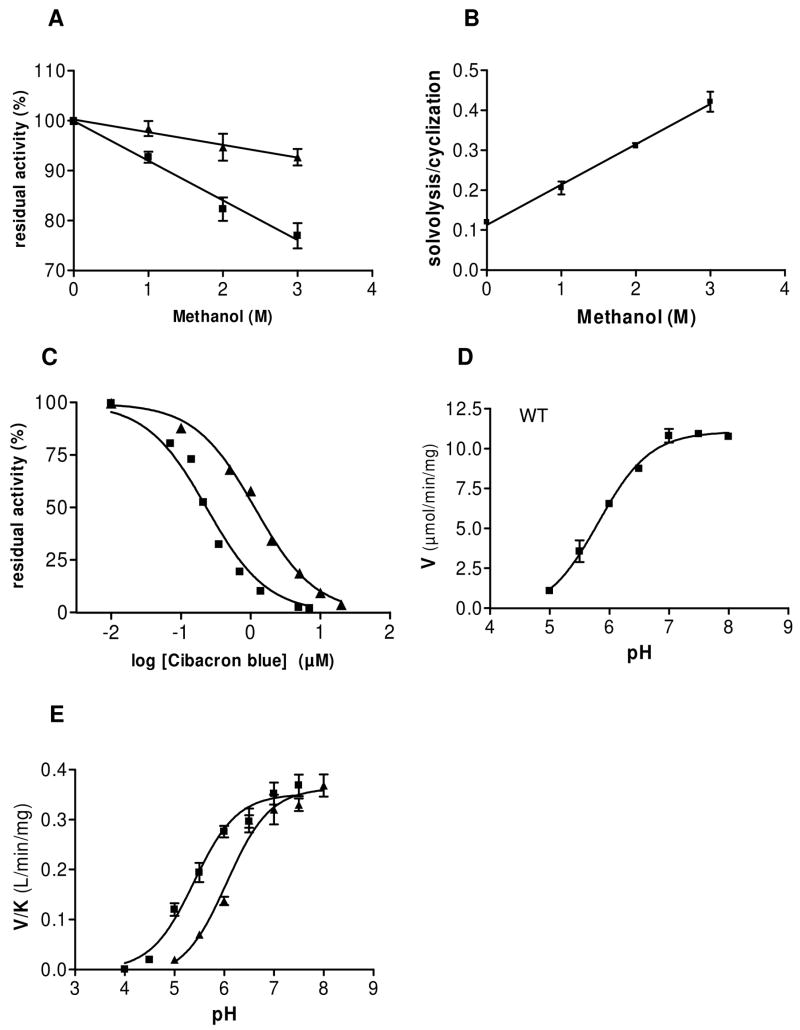
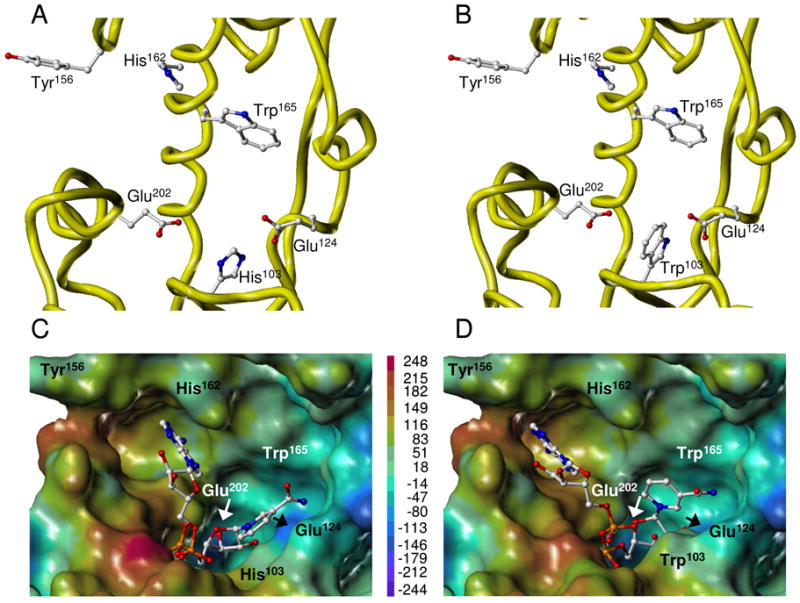
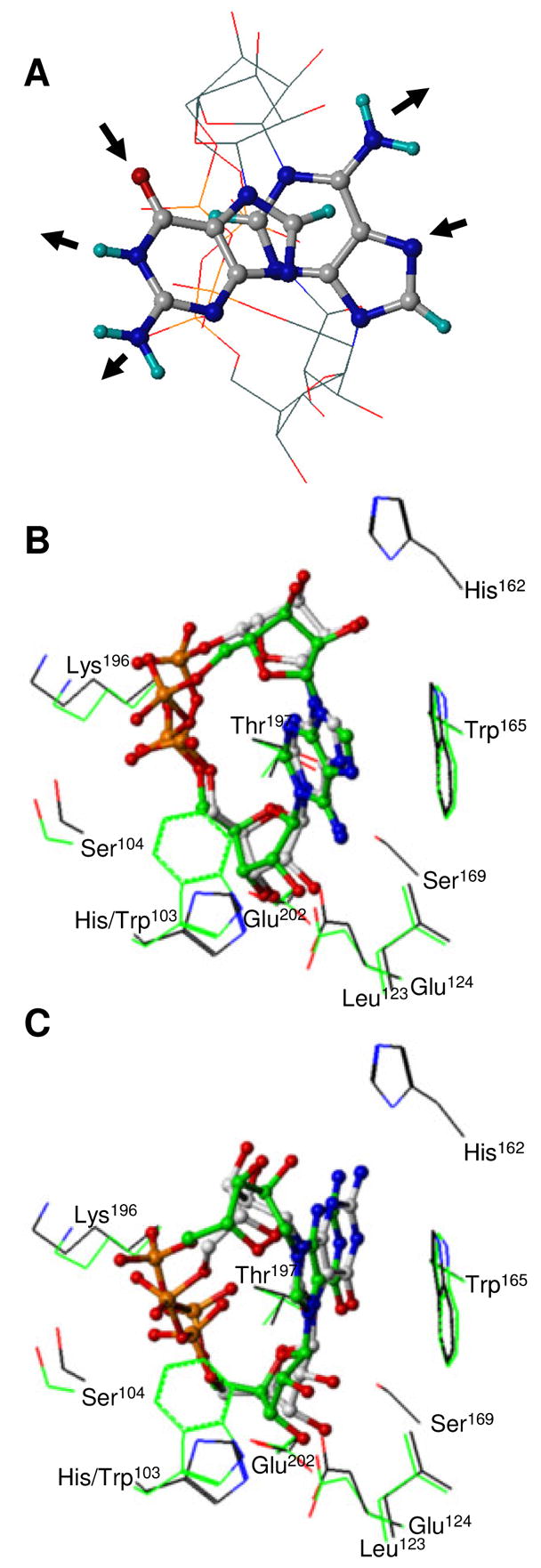
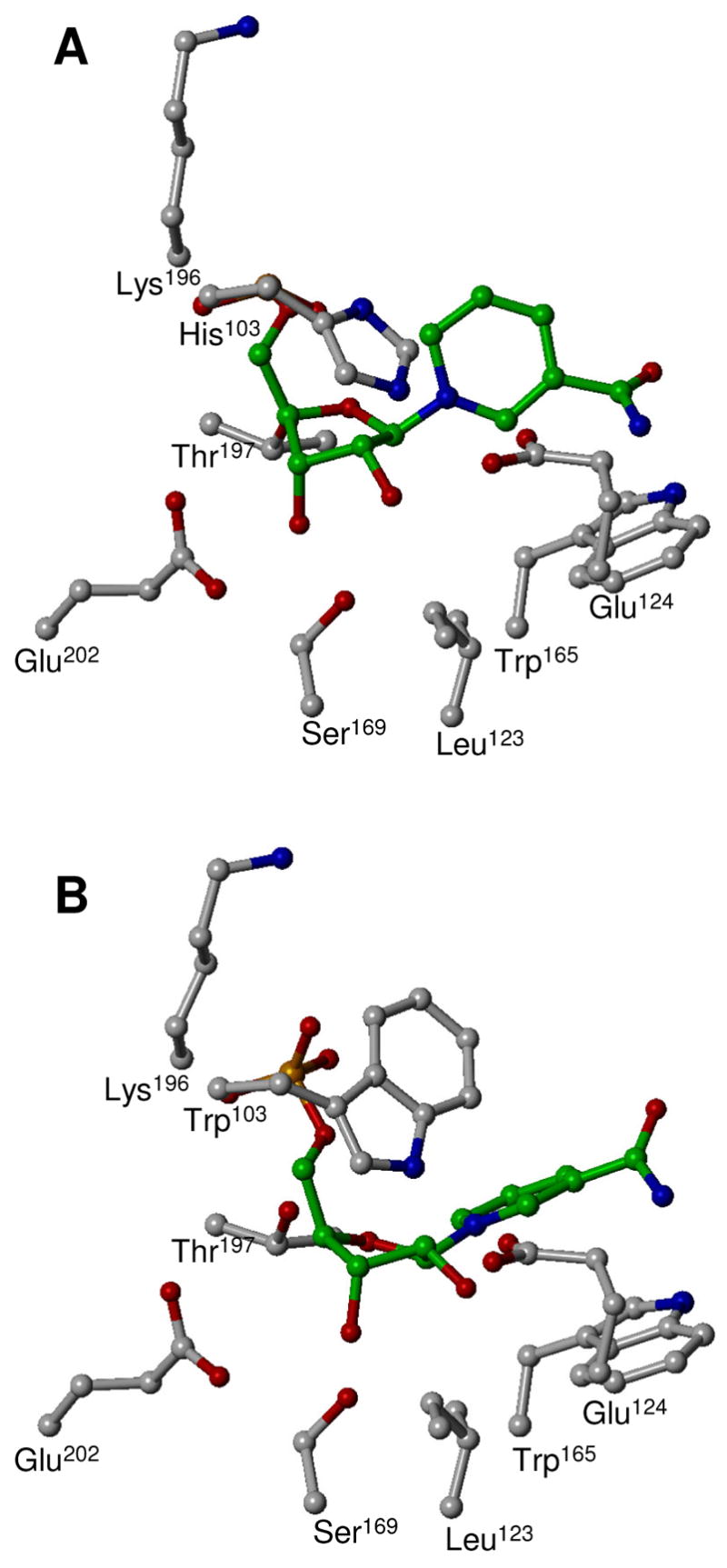
Schistosoma mansoni NAD(P)+ catabolizing enzyme (SmNACE) is a new member of the ADP-ribosyl cyclase family. In contrast to all the other enzymes that are involved in the production of metabolites that elicit Ca2+ mobilization, SmNACE is virtually unable to transform NAD+ into the second messenger cyclic ADP-ribose (cADPR). Sequence alignments revealed that one of four conserved residues within the active site of these enzymes was replaced in SmNACE by a histidine (His103) instead of the highly conserved tryptophan. To find out whether the inability of SmNACE to catalyze the canonical ADP-ribosyl cyclase reaction is linked to this change, we have replaced His103 with a tryptophan. The H103W mutation in SmNACE was indeed found to restore ADP-ribosyl cyclase activity as cADPR amounts for 7% of the reaction products (i.e., a value larger than observed for other members of this family such as CD38). Introduction of a Trp103 residue provides some of the binding characteristics of mammalian ADP-ribosyl cyclases such as increased affinity for Cibacron blue and slow-binding inhibition by araF-NAD+. Homology modeling of wild-type and H103W mutant three-dimensional structures, and docking of substrates within the active sites, provides new insight into the catalytic mechanism of SmNACE. Both residue side chains share similar roles in the nicotinamide-ribose bond cleavage step leading to an E.ADP-ribosyl reaction intermediate. They diverge, however, in the evolution of this intermediate; His103 provides a more polar environment favoring the accessibility to water and hydrolysis leading to ADP-ribose at the expense of the intramolecular cyclization pathway resulting in cADPR.
Figures

Similar articles
-
Schistosoma mansoni NAD(+) catabolizing enzyme: identification of key residues in catalysis.Biochim Biophys Acta. 2013 Dec;1834(12):2520-7. doi: 10.1016/j.bbapap.2013.09.002. Epub 2013 Sep 12. Biochim Biophys Acta. 2013. PMID: 24035885
-
Production of calcium-mobilizing metabolites by a novel member of the ADP-ribosyl cyclase family expressed in Schistosoma mansoni.Biochemistry. 2005 Aug 23;44(33):11082-97. doi: 10.1021/bi050704r. Biochemistry. 2005. PMID: 16101292
-
Mechanism of cyclizing NAD to cyclic ADP-ribose by ADP-ribosyl cyclase and CD38.J Biol Chem. 2009 Oct 2;284(40):27629-36. doi: 10.1074/jbc.M109.030965. Epub 2009 Jul 28. J Biol Chem. 2009. PMID: 19640843 Free PMC article.
-
The ADP-ribosyl cyclases--the current evolutionary state of the ARCs.Front Biosci (Landmark Ed). 2014 Jun 1;19(6):986-1002. doi: 10.2741/4262. Front Biosci (Landmark Ed). 2014. PMID: 24896331 Review.
-
Paracrine ADP Ribosyl Cyclase-Mediated Regulation of Biological Processes.Cells. 2022 Aug 24;11(17):2637. doi: 10.3390/cells11172637. Cells. 2022. PMID: 36078044 Free PMC article. Review.
Cited by
-
Identification of a major enzyme for the synthesis and hydrolysis of cyclic ADP-ribose in amphibian cells and evolutional conservation of the enzyme from human to invertebrate.Mol Cell Biochem. 2012 Jul;366(1-2):69-80. doi: 10.1007/s11010-012-1284-0. Epub 2012 Mar 16. Mol Cell Biochem. 2012. PMID: 22422046
-
Molecular characterization of a novel intracellular ADP-ribosyl cyclase.PLoS One. 2007 Aug 29;2(8):e797. doi: 10.1371/journal.pone.0000797. PLoS One. 2007. PMID: 17726527 Free PMC article.
-
Insights into the mechanism of bovine CD38/NAD+glycohydrolase from the X-ray structures of its Michaelis complex and covalently-trapped intermediates.PLoS One. 2012;7(4):e34918. doi: 10.1371/journal.pone.0034918. Epub 2012 Apr 18. PLoS One. 2012. PMID: 22529956 Free PMC article.
-
A single residue in a novel ADP-ribosyl cyclase controls production of the calcium-mobilizing messengers cyclic ADP-ribose and nicotinic acid adenine dinucleotide phosphate.J Biol Chem. 2010 Jun 25;285(26):19900-9. doi: 10.1074/jbc.M110.105312. Epub 2010 Apr 12. J Biol Chem. 2010. PMID: 20385553 Free PMC article.
References
-
- Howard M, Grimaldi JC, Bazan JF, Lund FE, Santos-Argumedo L, Parkhouse RM, Walseth TF, Lee HC. Formation and hydrolysis of cyclic ADP-ribose catalyzed by lymphocyte antigen CD38. Science. 1993;262:1056–1059. - PubMed
-
- Lee HC. Enzymatic functions and structures of CD38 and homologs. Chem Immunol. 2000;75:39–59. - PubMed
-
- Schuber F, Lund FE. Structure and enzymology of ADP-ribosyl cyclases: Conserved enzymes that produce multiple calcium mobilizing metabolites. Curr Mol Med. 2004;4:249–261. - PubMed
-
- Lee HC. Nicotinic acid adenine dinucleotide phosphate (NAADP)-mediated calcium signaling. J Biol Chem. 2005;280:33693–33696. - PubMed
-
- Hirata Y, Kimura N, Sato K, Ohsugi Y, Takasawa S, Okamoto H, Ishikawa J, Kaisho T, Ishihara K, Hirano T. ADP ribosyl cyclase activity of a novel bone marrow stromal cell surface molecule, BST-1. FEBS Lett. 1994;356:244–248. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
