

Evolution of new function in the GTP cyclohydrolase II proteins of Streptomyces coelicolor
- PMID: 17002314
- PMCID: PMC3227873
- DOI: 10.1021/bi061005x
Evolution of new function in the GTP cyclohydrolase II proteins of Streptomyces coelicolor
Abstract
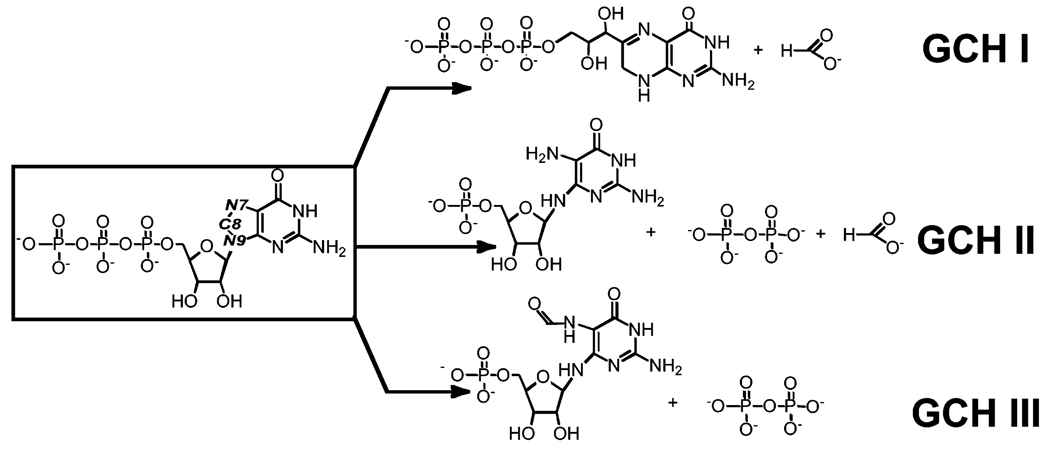
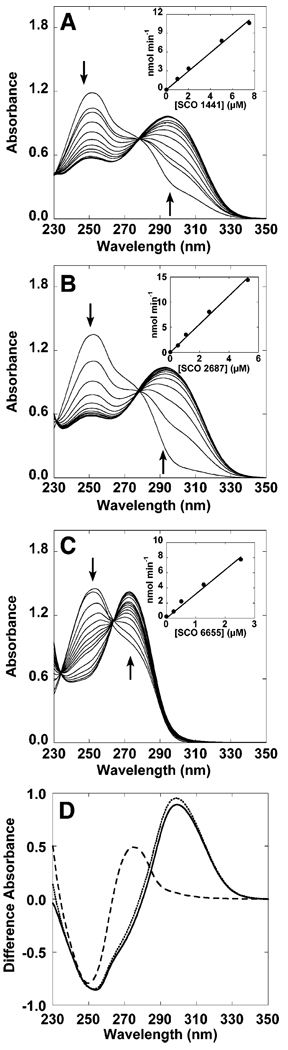
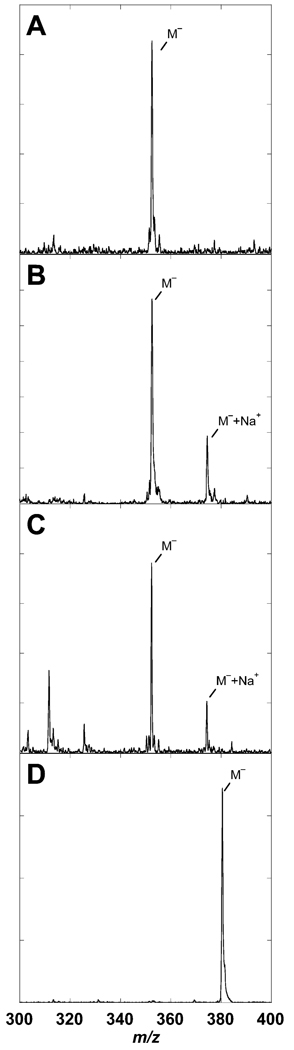
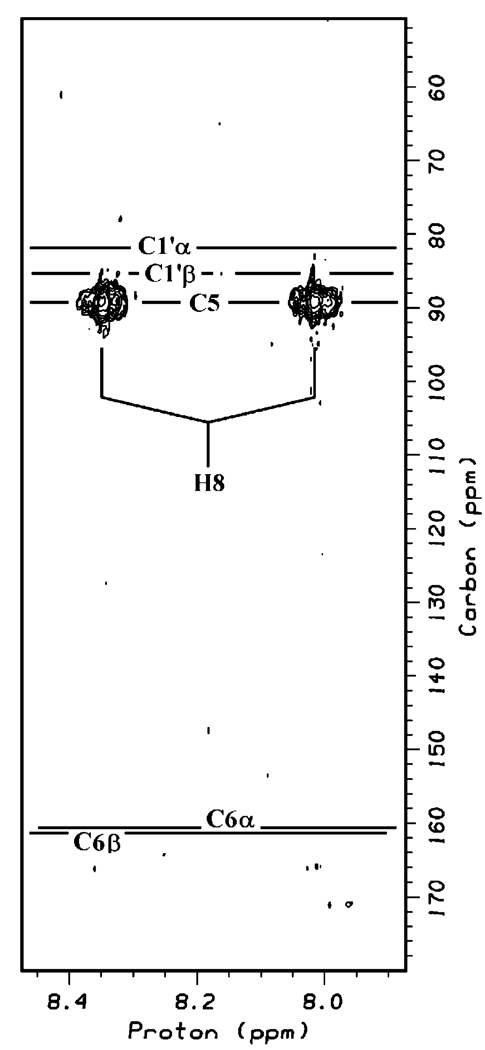
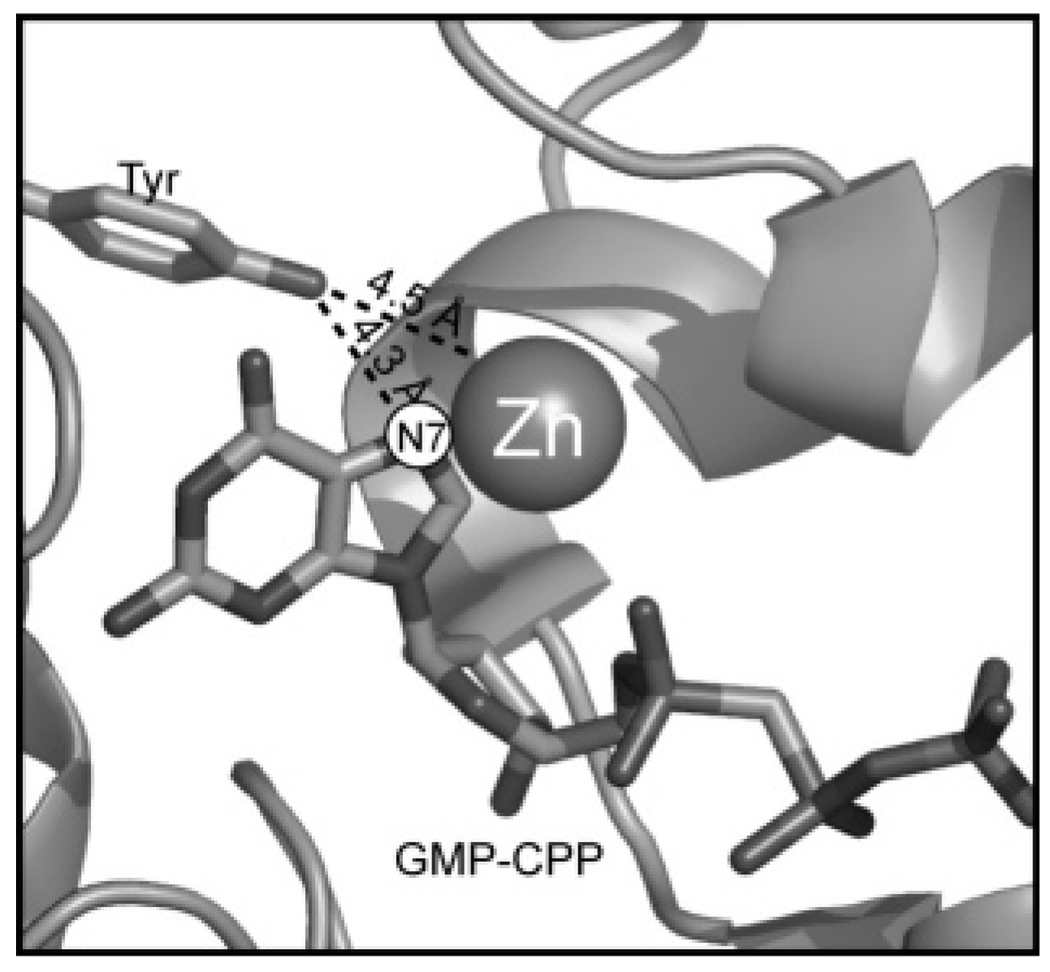
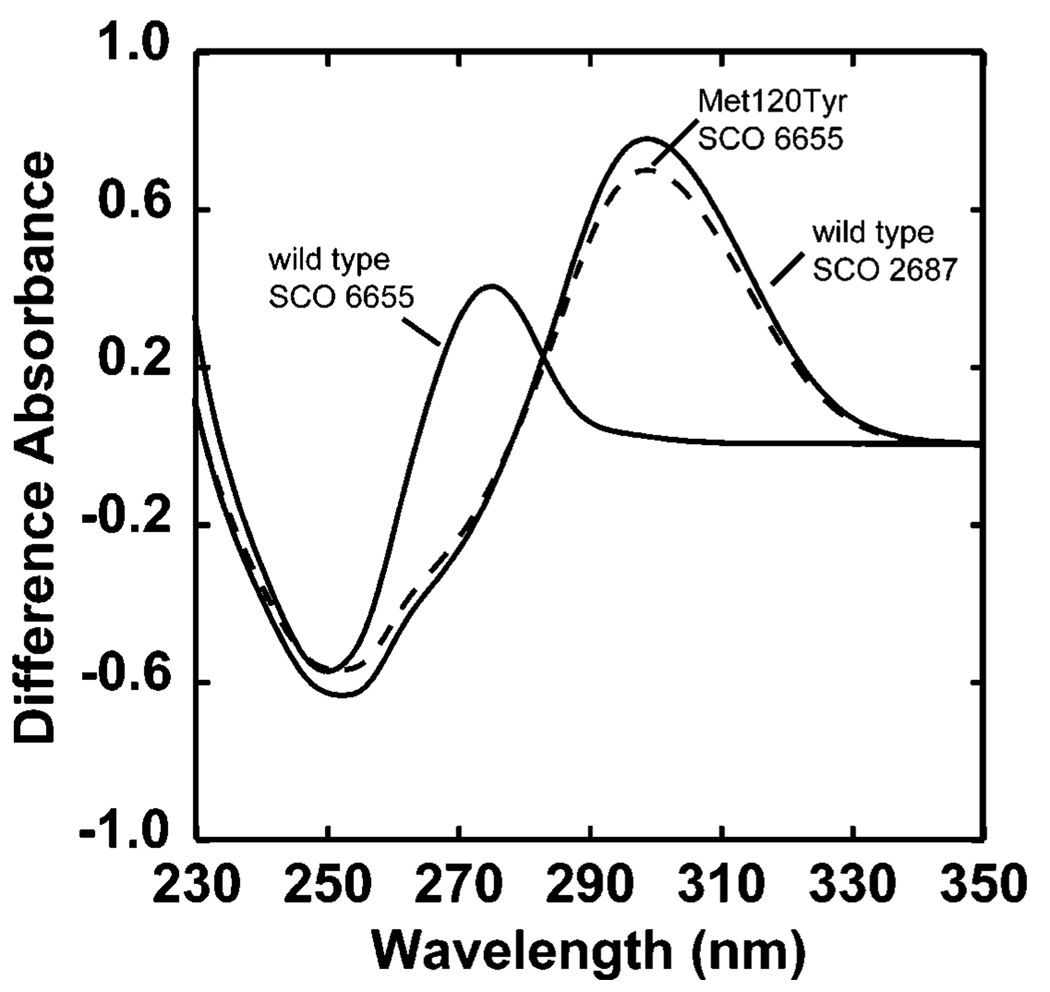
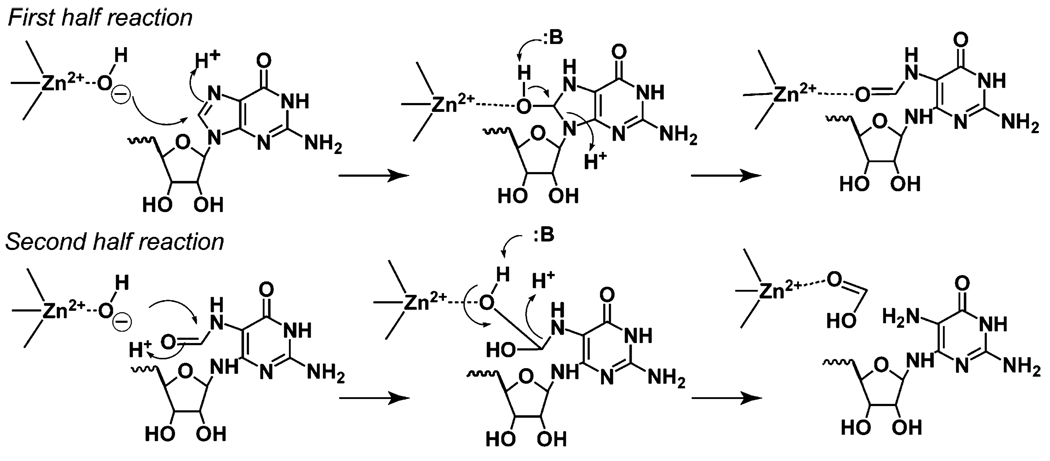
The genome sequence of Streptomyces coelicolor contains three open reading frames (sco1441, sco2687, and sco6655) that encode proteins with significant (>40%) amino acid identity to GTP cyclohydrolase II (GCH II), which catalyzes the committed step in the biosynthesis of riboflavin. The physiological significance of the redundancy of these proteins in S. coelicolor is not known. However, the gene contexts of the three proteins are different, suggesting that they may serve alternate biological niches. Each of the three proteins was overexpressed in Escherichia coli and characterized to determine if their functions are biologically overlapping. As purified, each protein contains 1 molar equiv of zinc/mol of protein and utilizes guanosine 5'-triphosphate (GTP) as substrate. Two of these proteins (SCO 1441 and SCO 2687) produce the canonical product of GCH II, 2,5-diamino-6-ribosylamino-4(3H)-pyrimidinone 5'-phosphate (APy). Remarkably, however, one of the three proteins (SCO 6655) converts GTP to 2-amino-5-formylamino-6-ribosylamino-4(3H)-pyrimidinone 5'-phosphate (FAPy), as shown by UV-visible spectrophotometry, mass spectrometry, and NMR. This activity has been reported for a GTP cyclohydrolase III protein from Methanocaldococcus jannaschii [Graham, D. E., Xu, H., and White, R. H. (2002) Biochemistry 41, 15074-15084], which has no amino acid sequence homology to SCO 6655. Comparison of the sequences of these proteins and mapping onto the structure of the E. coli GCH II protein [Ren, J., Kotaka, M., Lockyer, M., Lamb, H. K., Hawkins, A. R., and Stammers, D. K. (2005) J. Biol. Chem. 280, 36912-36919] allowed identification of a switch residue, Met120, which appears to be responsible for the altered fate of GTP observed with SCO 6655; a Tyr is found in the analogous position of all proteins that have been shown to catalyze the conversion of GTP to APy. The Met120Tyr variant of SCO 6655 acquires the ability to catalyze the conversion of GTP to APy, suggesting a role for Tyr120 in the late phase of the reaction. Our data are consistent with duplication of GCH II in S. coelicolor promoting evolution of a new function. The physiological role(s) of the gene clusters that house GCH II homologues will be discussed.
Figures

Similar articles
-
A new use for a familiar fold: the X-ray crystal structure of GTP-bound GTP cyclohydrolase III from Methanocaldococcus jannaschii reveals a two metal ion catalytic mechanism.Biochemistry. 2008 Jan 8;47(1):230-42. doi: 10.1021/bi701782e. Epub 2007 Dec 4. Biochemistry. 2008. PMID: 18052207
-
Understanding functional divergence in proteins by studying intragenomic homologues.Biochemistry. 2008 Feb 26;47(8):2592-600. doi: 10.1021/bi702263z. Biochemistry. 2008. PMID: 18281960
-
An iron(II) dependent formamide hydrolase catalyzes the second step in the archaeal biosynthetic pathway to riboflavin and 7,8-didemethyl-8-hydroxy-5-deazariboflavin.Biochemistry. 2009 May 19;48(19):4181-8. doi: 10.1021/bi802341p. Biochemistry. 2009. PMID: 19309161
-
Biosynthesis of vitamin b2 (riboflavin).Annu Rev Nutr. 2000;20:153-67. doi: 10.1146/annurev.nutr.20.1.153. Annu Rev Nutr. 2000. PMID: 10940330 Review.
-
GTP-cyclohydrolases: a review.J Clin Chem Clin Biochem. 1985 Apr;23(4):169-76. J Clin Chem Clin Biochem. 1985. PMID: 3891906 Review.
Cited by
-
Deciphering deazapurine biosynthesis: pathway for pyrrolopyrimidine nucleosides toyocamycin and sangivamycin.Chem Biol. 2008 Aug 25;15(8):790-8. doi: 10.1016/j.chembiol.2008.07.012. Chem Biol. 2008. PMID: 18721750 Free PMC article.
-
Genetic control of biosynthesis and transport of riboflavin and flavin nucleotides and construction of robust biotechnological producers.Microbiol Mol Biol Rev. 2011 Jun;75(2):321-60. doi: 10.1128/MMBR.00030-10. Microbiol Mol Biol Rev. 2011. PMID: 21646432 Free PMC article. Review.
-
Arabidopsis RIBA proteins: two out of three isoforms have lost their bifunctional activity in riboflavin biosynthesis.Int J Mol Sci. 2012 Oct 31;13(11):14086-105. doi: 10.3390/ijms131114086. Int J Mol Sci. 2012. PMID: 23203051 Free PMC article.
-
Structure of a 6-pyruvoyltetrahydropterin synthase homolog from Streptomyces coelicolor.Acta Crystallogr Sect F Struct Biol Cryst Commun. 2008 Oct 1;64(Pt 10):875-9. doi: 10.1107/S1744309108027048. Epub 2008 Sep 30. Acta Crystallogr Sect F Struct Biol Cryst Commun. 2008. PMID: 18931427 Free PMC article.
References
-
- Ohno S. Evolution by gene duplication. New York: Springer-Verlag; 1970.
-
- Jensen RA. Enzyme recruitiment in evolution of new function. Annu. Rev. Microbiol. 1976;30:409–425. - PubMed
-
- Bentley SD, Chater KF, Cerdeno-Tarraga AM, Challis GL, Thomson NR, James KD, Harris DE, Quail MA, Kieser H, Harper D, Bateman A, Brown S, Chandra G, Chen CW, Collins M, Cronin A, Fraser A, Goble A, Hidalgo J, Hornsby T, Howarth S, Huang CH, Kieser T, Larke L, Murphy L, Oliver K, O’Neil S, Rabbinowitsch E, Rajandream MA, Rutherford K, Rutter S, Seeger K, Saunders D, Sharp S, Squares R, Squares S, Taylor K, Warren T, Wietzorrek A, Woodward J, Barrell BG, Parkhill J, Hopwood DA. Complete genome sequence of the model actinomycete Streptomyces coelicolor A3(2) Nature. 2002;417:141–147. - PubMed
-
- Gevers D, Vandepoele K, Simillon C, Van de Peer Y. Gene duplication and biased functional retention of paralogs in bacterial genomes. Trends Microbiol. 2004;12:148–154. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
