

The Rx-like homeobox gene (Rx-L) is necessary for normal photoreceptor development
- PMID: 17003412
- PMCID: PMC2824340
- DOI: 10.1167/iovs.06-0167
The Rx-like homeobox gene (Rx-L) is necessary for normal photoreceptor development
Abstract
Purpose: The retinal homeobox (Rx) gene plays an essential role in retinal development. An Rx-like (Rx-L) gene from Xenopus laevis has been identified. The purpose of this study was to analyze the function of Rx-L in the developing retina.
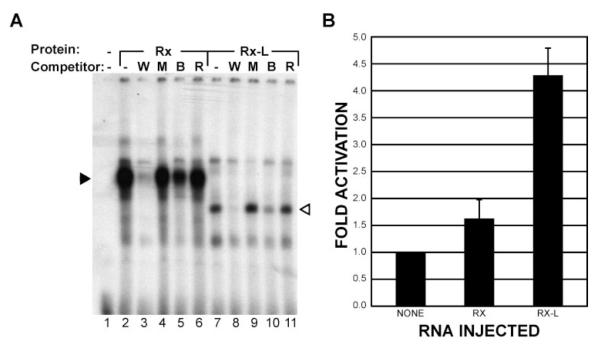
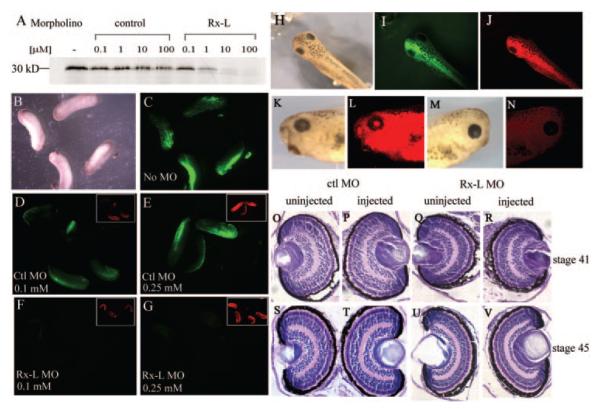
Methods: DNA-binding properties of Rx-L were analyzed by electrophoretic mobility shift assay (EMSA), with in vitro-translated proteins and radiolabeled oligonucleotide probe. The Rx-L expression pattern was analyzed by in situ hybridization using whole or sectioned embryos and digoxigenin-labeled antisense riboprobes. Rx-L loss of function was studied by using antisense morpholino oligonucleotides targeted to the Rx-L translation initiation site. Embryos injected with control or Rx-L morpholinos were analyzed at stage 41 or 45.
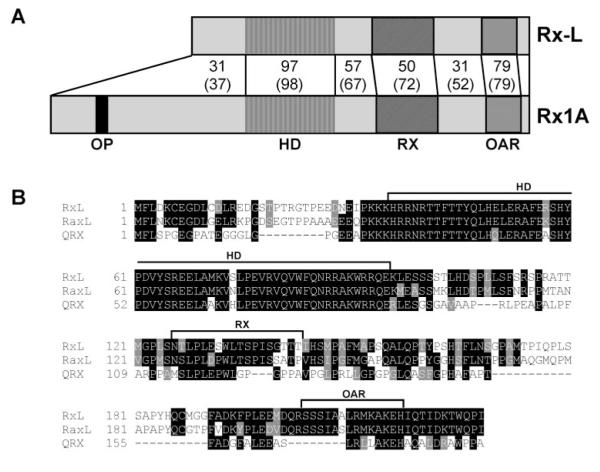
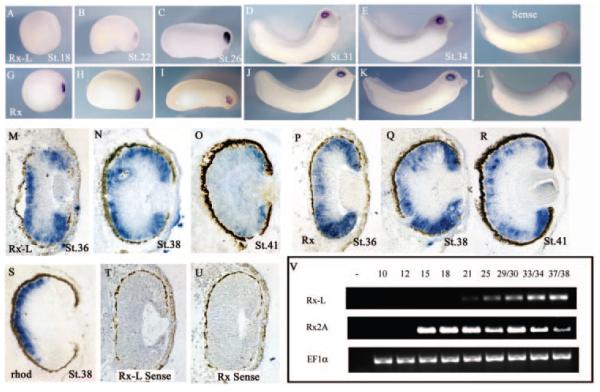
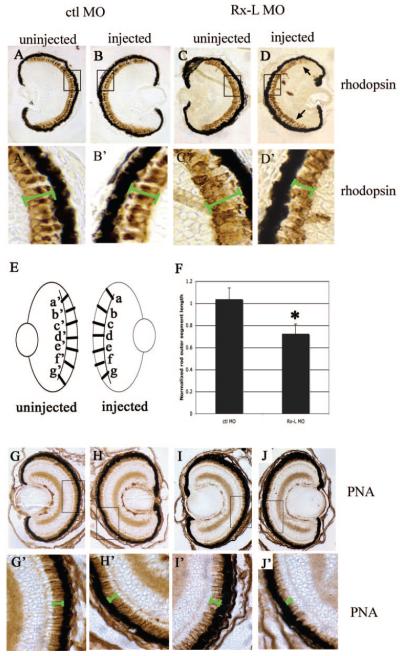
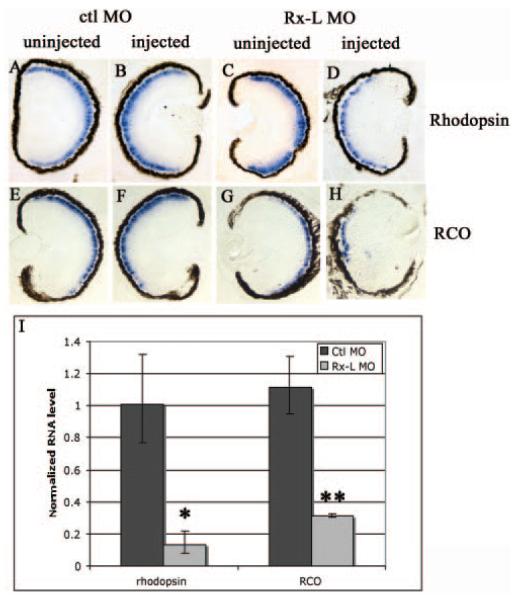
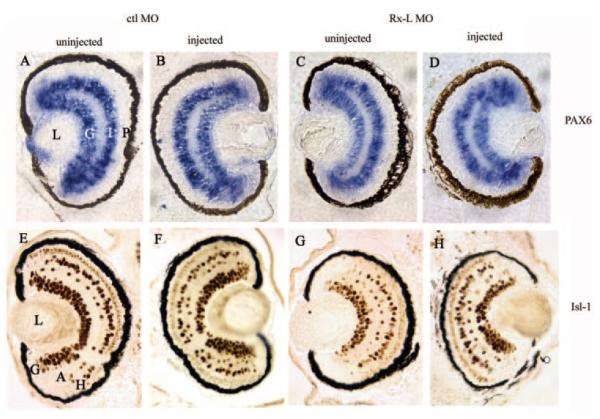
Results: Rx-L shares homology with Rx at the homeo-, OAR, and Rx domains, but lacks an octapeptide motif. Rx-L is expressed in the developing retina beginning in the early tailbud stage. In the maturing retina, Rx-L expression is restricted primarily to the developing photoreceptor layer and the ciliary marginal zone. Rx-L can bind a photoreceptor conserved element-1 (PCE-1) oligonucleotide, an element conserved among all known photoreceptor gene promoters. In a promoter activity assay, Rx-L functions as a stronger transcriptional activator than Rx. Antisense morpholino-mediated knockdown of Rx-L expression resulted in a decrease in rhodopsin and red cone opsin expression levels in Xenopus retinas. Injection of the Rx-L antisense morpholino oligonucleotide also resulted in a decrease in the length of both rod and cone outer segments.
Conclusions: The results suggest that Rx-L functions to regulate rod and cone development by activating photoreceptor-specific gene expression.
Figures
Similar articles
-
Regulation of photoreceptor gene expression by the retinal homeobox (Rx) gene product.Dev Biol. 2010 Mar 15;339(2):494-506. doi: 10.1016/j.ydbio.2009.12.032. Epub 2010 Jan 7. Dev Biol. 2010. PMID: 20060393 Free PMC article.
-
Regulation of photoreceptor gene transcription via a highly conserved transcriptional regulatory element by vsx gene products.Mol Vis. 2016 Dec 14;22:1421-1428. eCollection 2016. Mol Vis. 2016. PMID: 28003732 Free PMC article.
-
Delayed expression of the Crx gene and photoreceptor development in the Chx10-deficient retina.Invest Ophthalmol Vis Sci. 2004 Feb;45(2):375-84. doi: 10.1167/iovs.03-0332. Invest Ophthalmol Vis Sci. 2004. PMID: 14744875
-
Regulation of vertebrate eye development by Rx genes.Int J Dev Biol. 2004;48(8-9):761-70. doi: 10.1387/ijdb.041878tb. Int J Dev Biol. 2004. PMID: 15558469 Review.
-
Expression of FoxE4 and Rx visualizes the timing and dynamics of critical processes taking place during initial stages of vertebrate eye development.Dev Neurosci. 2004;26(5-6):294-307. doi: 10.1159/000082271. Dev Neurosci. 2004. PMID: 15855758 Review.
Cited by
-
Xenopus mutant reveals necessity of rax for specifying the eye field which otherwise forms tissue with telencephalic and diencephalic character.Dev Biol. 2014 Nov 15;395(2):317-330. doi: 10.1016/j.ydbio.2014.09.004. Epub 2014 Sep 16. Dev Biol. 2014. PMID: 25224223 Free PMC article.
-
Regulation of retinal homeobox gene transcription by cooperative activity among cis-elements.Gene. 2010 Nov 1;467(1-2):13-24. doi: 10.1016/j.gene.2010.07.005. Epub 2010 Jul 11. Gene. 2010. PMID: 20627122 Free PMC article.
-
The rax homeobox gene is mutated in the eyeless axolotl, Ambystoma mexicanum.Dev Dyn. 2021 Jun;250(6):807-821. doi: 10.1002/dvdy.246. Epub 2020 Sep 17. Dev Dyn. 2021. PMID: 32864847 Free PMC article.
-
Regulation of photoreceptor gene expression by the retinal homeobox (Rx) gene product.Dev Biol. 2010 Mar 15;339(2):494-506. doi: 10.1016/j.ydbio.2009.12.032. Epub 2010 Jan 7. Dev Biol. 2010. PMID: 20060393 Free PMC article.
-
Eye evolution: common use and independent recruitment of genetic components.Philos Trans R Soc Lond B Biol Sci. 2009 Oct 12;364(1531):2819-32. doi: 10.1098/rstb.2009.0079. Philos Trans R Soc Lond B Biol Sci. 2009. PMID: 19720647 Free PMC article. Review.
References
-
- Zaghloul NA, Yan B, Moody SA. Step-wise specification of retinal stem cells during normal embryogenesis. Biol Cell. 2005;97:321–337. - PubMed
-
- Chow RL, Lang RA. Early eye development in vertebrates. Annu Rev Cell Dev Biol. 2001;17:255–296. - PubMed
-
- Marquardt T, Gruss P. Generating neuronal diversity in the retina: one for nearly all. Trends Neurosci. 2002;25:32–38. - PubMed
-
- Marquardt T. Transcriptional control of neuronal diversification in the retina. Prog Retin Eye Res. 2003;22:567–577. - PubMed
-
- Zuber ME, Gestri G, Viczian AS, Barsacchi G, Harris WA. Specification of the vertebrate eye by a network of eye field transcription factors. Development. 2003;130:5155–5167. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
