

Differential enhancement of dengue virus immune complex infectivity mediated by signaling-competent and signaling-incompetent human Fcgamma RIA (CD64) or FcgammaRIIA (CD32)
- PMID: 17005690
- PMCID: PMC1617290
- DOI: 10.1128/JVI.00792-06
Differential enhancement of dengue virus immune complex infectivity mediated by signaling-competent and signaling-incompetent human Fcgamma RIA (CD64) or FcgammaRIIA (CD32)
Abstract
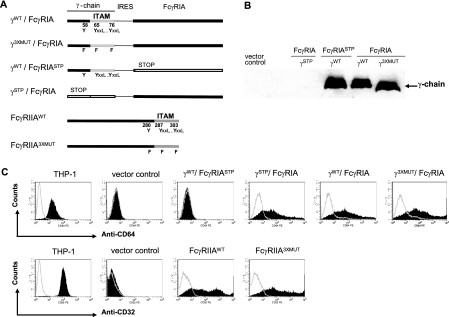
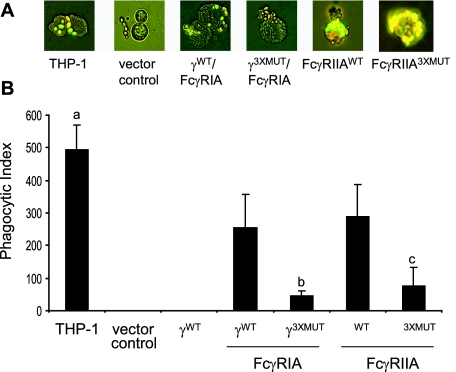
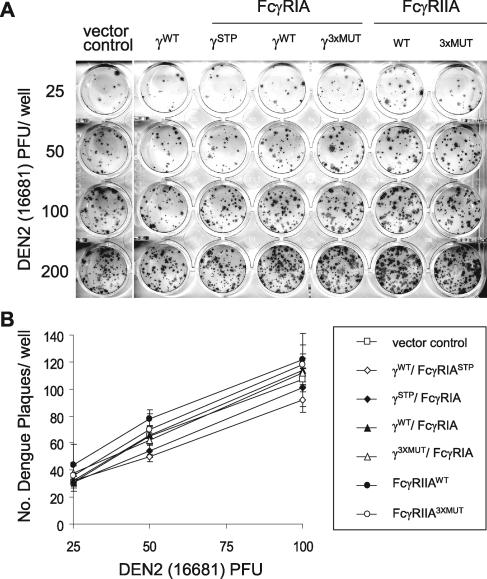
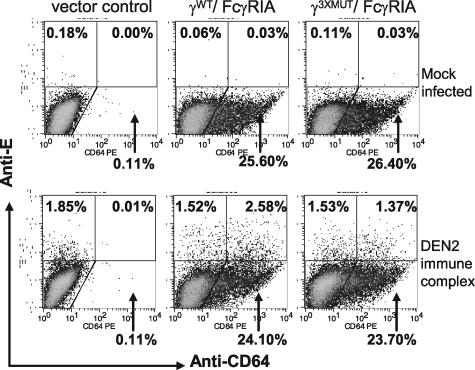
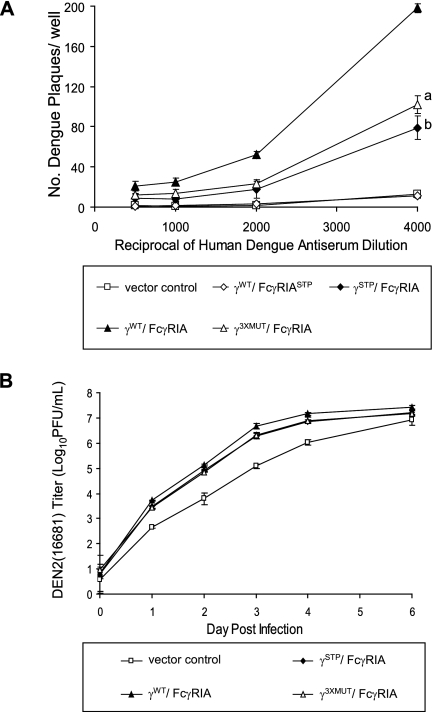
Fcgamma receptor (FcgammaR)-mediated entry of infectious dengue virus immune complexes into monocytes/macrophages is hypothesized to be a key event in the pathogenesis of complicated dengue fever. FcgammaRIA (CD64) and FcgammaRIIA (CD32), which predominate on the surface of such dengue virus-permissive cells, were compared for their influence on the infectivity of dengue 2 virus immune complexes formed with human dengue virus antibodies. A signaling immunoreceptor tyrosine-based activation motif (ITAM) incorporated into the accessory gamma-chain subunit that associates with FcgammaRIA and constitutively in FcgammaRIIA is required for phagocytosis mediated by these receptors. To determine whether FcgammaRIA and FcgammaRIIA activation functions are also required for internalization of infectious dengue virus immune complexes, we generated native and signaling-incompetent versions of each receptor by site-directed mutagenesis of ITAM tyrosine residues. Plasmids designed to express these receptors were transfected into COS-7 cells, and dengue virus replication was measured by plaque assay and flow cytometry. We found that both receptors mediated enhanced dengue virus immune complex infectivity but that FcgammaRIIA appeared to do so far more effectively. Abrogation of FcgammaRIA signaling competency, either by expression without gamma-chain or by coexpression with gamma-chain mutants, was associated with significant impairment of phagocytosis and of dengue virus immune complex infectivity. Abrogation of FcgammaRIIA signaling competency was also associated with equally impaired phagocytosis but had no discernible effect on dengue virus immune complex infectivity. These findings point to fundamental differences between FcgammaRIA and FcgammaRIIA with respect to their immune-enhancing capabilities and suggest that different mechanisms of dengue virus immune complex internalization may operate between these FcgammaRs.
Figures
References
-
- Aderem, A., and D. M. Underhill. 1999. Mechanisms of phagocytosis in macrophages. Annu. Rev. Immunol. 17:593-623. - PubMed
-
- Daeron, M. 1997. Fc receptor biology. Annu. Rev. Immunol. 15:203-234. - PubMed
-
- Duchemin, A. M., L. K. Ernst, and C. L. Anderson. 1994. Clustering of the high affinity Fc receptor for immunoglobulin G (Fc gamma RI) results in phosphorylation of its associated gamma-chain. J. Biol. Chem. 269:12111-12117. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
