

Local interneurons regulate synaptic strength by retrograde release of endocannabinoids
- PMID: 17005857
- PMCID: PMC6674464
- DOI: 10.1523/JNEUROSCI.0958-06.2006
Local interneurons regulate synaptic strength by retrograde release of endocannabinoids
Abstract
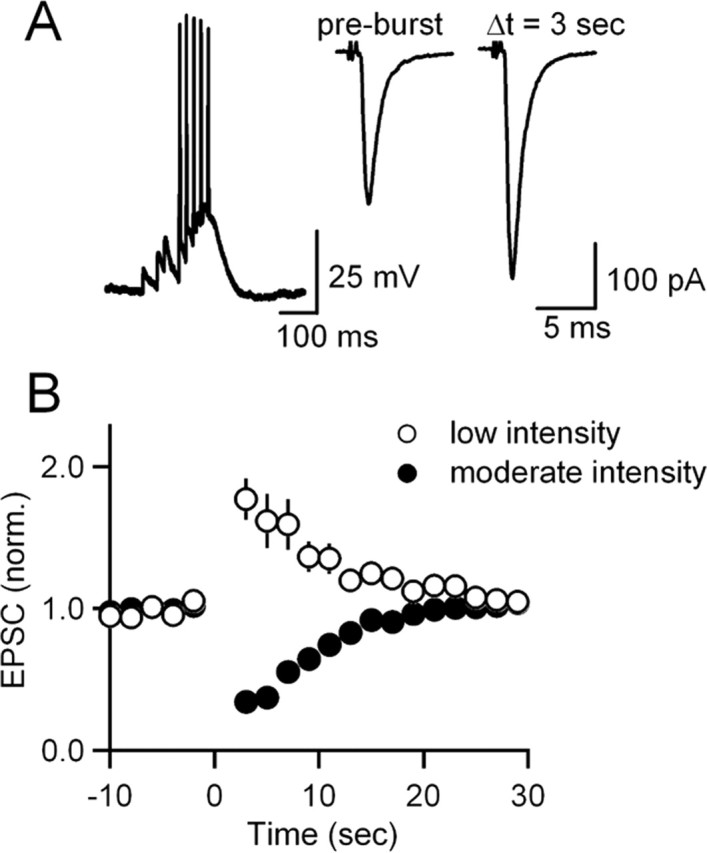
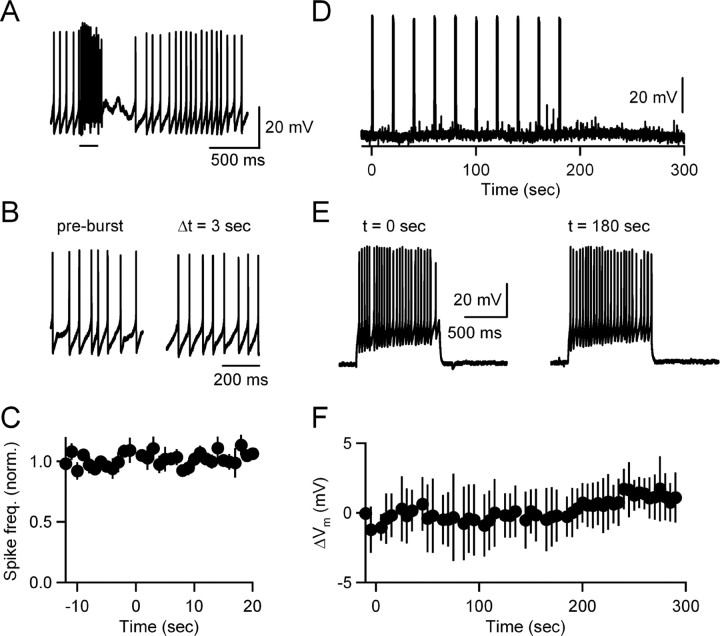
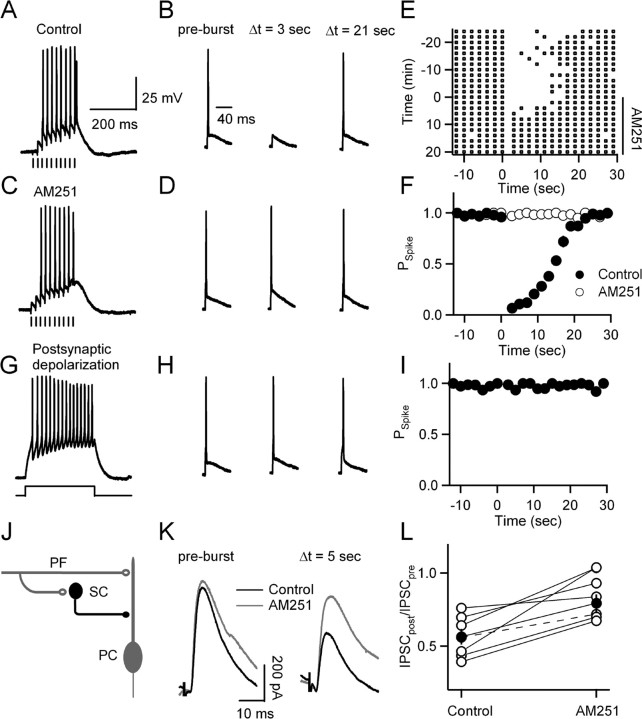
Neurons release endocannabinoids from their dendrites to trigger changes in the probability of transmitter release. Although such retrograde signaling has been described for principal neurons, such as hippocampal pyramidal cells and cerebellar Purkinje cells (PCs), it has not been demonstrated for local interneurons. Here we tested whether inhibitory interneurons in the cerebellum, stellate cells (SCs) and basket cells, regulate the strength of parallel fiber (PF) synapses by releasing endocannabinoids. We found that depolarization-induced suppression of excitation (DSE) is present in both SCs and basket cells. The properties of retrograde inhibition were examined more thoroughly for SCs. Both DSE and synaptically evoked suppression of excitation (SSE) triggered with brief PF bursts require elevations of postsynaptic calcium, are blocked by a type 1 cannabinoid receptor (CB1R) antagonist, and are absent in mice lacking the CB1R. SSE for SCs is similar to that described previously for PCs in that it is prevented by BAPTA and DAG lipase inhibitors in the recording pipette; however, unlike in PCs, NMDA receptors (NMDARs) play an important role in SSE for SCs. Although SCs express CB1Rs postsynaptically, neither high-frequency firing of SCs nor PF bursts lead to autocrine suppression of subsequent SC activity. Instead, PF bursts decrease the amplitude of disynaptic inhibition in PCs by evoking endocannabinoid release that transiently reduces the ability of PF synapses to trigger spikes in SCs. Thus, local interneurons within the cerebellum can release endocannabinoids through metabotropic glutamate receptor- and NMDAR-dependent mechanisms and contribute to use-dependent modulation of circuit properties.
Figures
Comment in
-
Endocannabinoid-dependent regulation of feedforward inhibition in cerebellar Purkinje cells.J Neurosci. 2007 Jan 3;27(1):1-3. doi: 10.1523/jneurosci.4842-06.2007. J Neurosci. 2007. PMID: 17205618 Free PMC article. Review. No abstract available.
References
-
- Bacci A, Huguenard JR, Prince DA. Long-lasting self-inhibition of neocortical interneurons mediated by endocannabinoids. Nature. 2004;431:312–316. - PubMed
-
- Bisogno T, Howell F, Williams G, Minassi A, Cascio MG, Ligresti A, Matias I, Schiano-Moriello A, Paul P, Williams EJ, Gangadharan U, Hobbs C, Di Marzo V, Doherty P. Cloning of the first sn1-DAG lipases points to the spatial and temporal regulation of endocannabinoid signaling in the brain. J Cell Biol. 2003;163:463–468. - PMC - PubMed
-
- Bisogno T, Ligresti A, Di Marzo V. The endocannabinoid signaling system: biochemical aspects. Pharmacol Biochem Behav. 2005;81:224–238. - PubMed
-
- Blitz DM, Regehr WG. Timing and specificity of feed-forward inhibition within the LGN. Neuron. 2005;45:917–928. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources