

TIF1 activates the intra-S-phase checkpoint response in the diploid micronucleus and amitotic polyploid macronucleus of Tetrahymena
- PMID: 17005912
- PMCID: PMC1679683
- DOI: 10.1091/mbc.e06-05-0469
TIF1 activates the intra-S-phase checkpoint response in the diploid micronucleus and amitotic polyploid macronucleus of Tetrahymena
Abstract
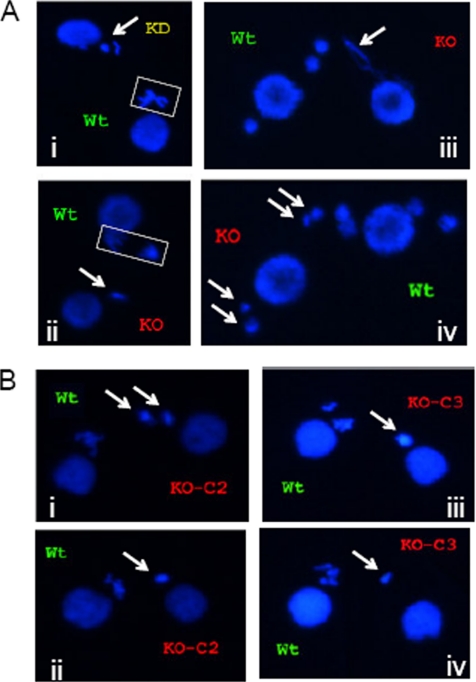
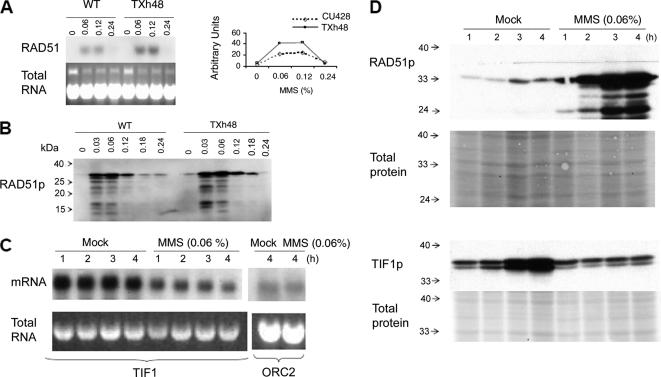
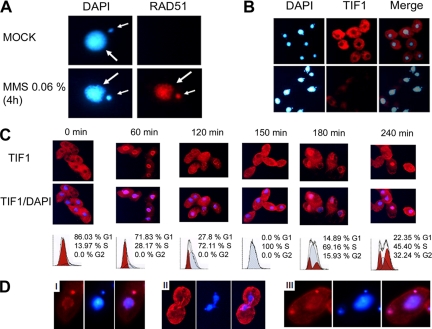
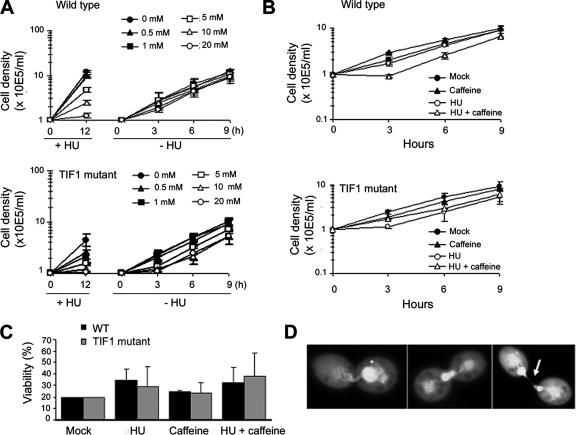
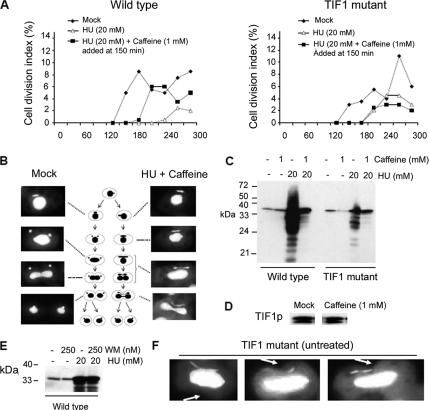
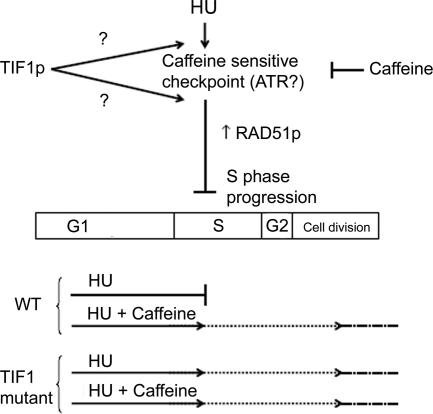
The ribosomal DNA origin binding protein Tif1p regulates the timing of rDNA replication and is required globally for proper S-phase progression and division of the Tetrahymena thermophila macronucleus. Here, we show that Tif1p safeguards chromosomes from DNA damage in the mitotic micronucleus and amitotic macronucleus. TIF1p localization is dynamically regulated as it moves into the micro- and macronucleus during the respective S phases. TIF1 disruption mutants are hypersensitive to hydroxyurea and methylmethanesulfonate, inducers of DNA damage and intra-S-phase checkpoint arrest in all examined eukaryotes. TIF1 mutants incur double-strand breaks in the absence of exogenous genotoxic stress, destabilizing all five micronuclear chromosomes. Wild-type Tetrahymena elicits an intra-S-phase checkpoint response that is induced by hydroxyurea and suppressed by caffeine, an inhibitor of the apical checkpoint kinase ATR/MEC1. In contrast, hydroxyurea-challenged TIF1 mutants fail to arrest in S phase or exhibit caffeine-sensitive Rad51 overexpression, indicating the involvement of TIF1 in checkpoint activation. Although aberrant micro- and macronuclear division occurs in TIF1 mutants and caffeine-treated wild-type cells, TIF1p bears no similarity to ATR or its substrates. We propose that TIF1 and ATR function in the same epistatic pathway to regulate checkpoint responses in the diploid mitotic micronucleus and polyploid amitotic macronucleus.
Figures

Similar articles
-
TIF1 Represses rDNA replication initiation, but promotes normal S phase progression and chromosome transmission in Tetrahymena.Mol Biol Cell. 2005 Jun;16(6):2624-35. doi: 10.1091/mbc.e05-02-0107. Epub 2005 Mar 16. Mol Biol Cell. 2005. PMID: 15772155 Free PMC article.
-
Phosphorylation of the SQ H2A.X motif is required for proper meiosis and mitosis in Tetrahymena thermophila.Mol Cell Biol. 2007 Apr;27(7):2648-60. doi: 10.1128/MCB.01910-06. Epub 2007 Jan 22. Mol Cell Biol. 2007. PMID: 17242195 Free PMC article.
-
Tetrahymena meiotic nuclear reorganization is induced by a checkpoint kinase-dependent response to DNA damage.Mol Biol Cell. 2009 May;20(9):2428-37. doi: 10.1091/mbc.e08-10-1058. Epub 2009 Mar 18. Mol Biol Cell. 2009. PMID: 19297526 Free PMC article.
-
Developmental progression of Tetrahymena through the cell cycle and conjugation.Methods Cell Biol. 2012;109:177-236. doi: 10.1016/B978-0-12-385967-9.00007-4. Methods Cell Biol. 2012. PMID: 22444146 Review.
-
Keeping the soma free of transposons: programmed DNA elimination in ciliates.J Biol Chem. 2011 Oct 28;286(43):37045-52. doi: 10.1074/jbc.R111.276964. Epub 2011 Sep 13. J Biol Chem. 2011. PMID: 21914793 Free PMC article. Review.
Cited by
-
Checkpoint Activation of an Unconventional DNA Replication Program in Tetrahymena.PLoS Genet. 2015 Jul 28;11(7):e1005405. doi: 10.1371/journal.pgen.1005405. eCollection 2015 Jul. PLoS Genet. 2015. PMID: 26218270 Free PMC article.
-
MRE11 and COM1/SAE2 are required for double-strand break repair and efficient chromosome pairing during meiosis of the protist Tetrahymena.Chromosoma. 2010 Oct;119(5):505-18. doi: 10.1007/s00412-010-0274-9. Epub 2010 Apr 27. Chromosoma. 2010. PMID: 20422424
-
Differential targeting of Tetrahymena ORC to ribosomal DNA and non-rDNA replication origins.EMBO J. 2009 Feb 4;28(3):223-33. doi: 10.1038/emboj.2008.282. Epub 2009 Jan 15. EMBO J. 2009. PMID: 19153611 Free PMC article.
-
Crystal structures of DNA-Whirly complexes and their role in Arabidopsis organelle genome repair.Plant Cell. 2010 Jun;22(6):1849-67. doi: 10.1105/tpc.109.071399. Epub 2010 Jun 15. Plant Cell. 2010. PMID: 20551348 Free PMC article.
-
An Optimized and Versatile Counter-Flow Centrifugal Elutriation Workflow to Obtain Synchronized Eukaryotic Cells.Front Cell Dev Biol. 2021 Apr 20;9:664418. doi: 10.3389/fcell.2021.664418. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 33959616 Free PMC article.
References
-
- Abraham R. T. Cell cycle checkpoint signaling through the ATM and ATR kinases. Genes Dev. 2001;15:2177–2196. - PubMed
-
- Allen S. L. Genomic exclusion–a rapid means for inducing homozygous diploid lines in Tetrahymena pyriformis syngen I. Science. 1967;155:575–577. - PubMed
-
- Bodenbender J., Prohaska A., Jauker F., Hipke H., Cleffmann G. DNA elimination and its relation to quantities in the macronucleus of Tetrahymena. Dev. Genet. 1992;13:103–110. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
