

The Dbs PH domain contributes independently to membrane targeting and regulation of guanine nucleotide-exchange activity
- PMID: 17007612
- PMCID: PMC1698603
- DOI: 10.1042/BJ20061020
The Dbs PH domain contributes independently to membrane targeting and regulation of guanine nucleotide-exchange activity
Abstract
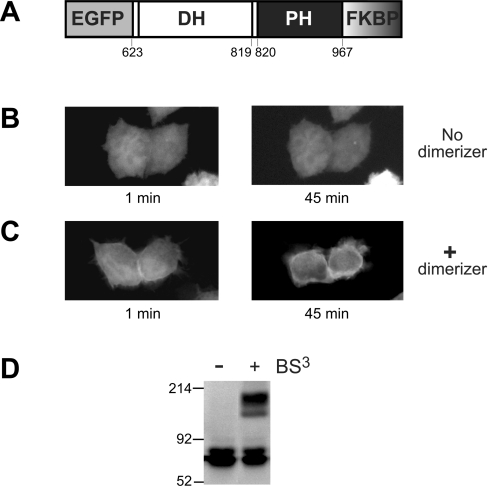
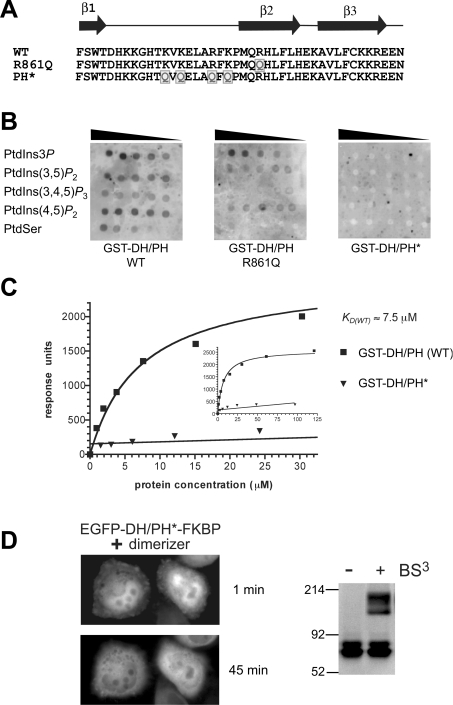
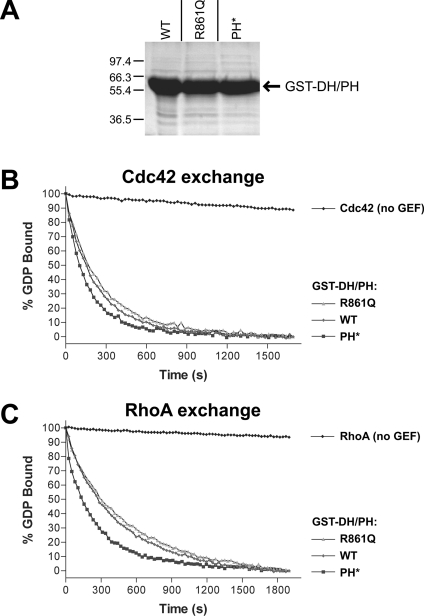
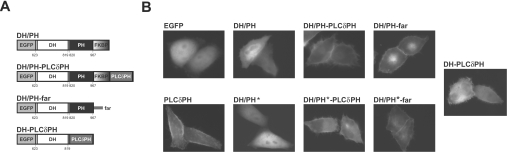
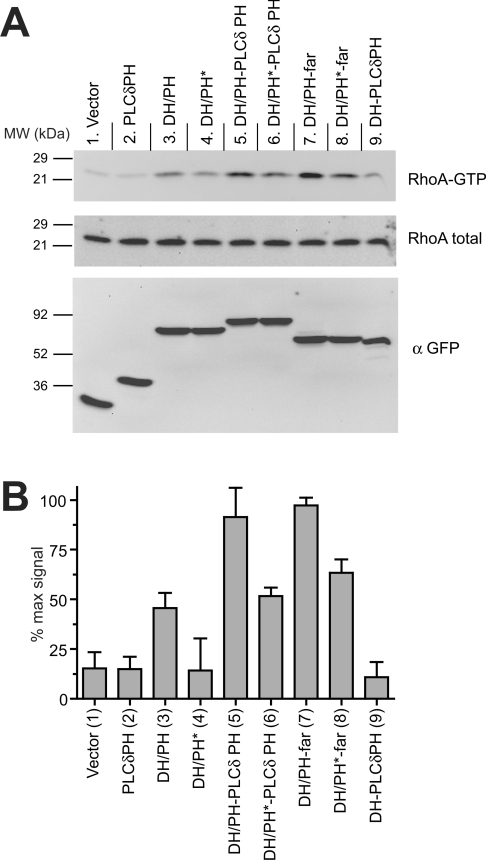
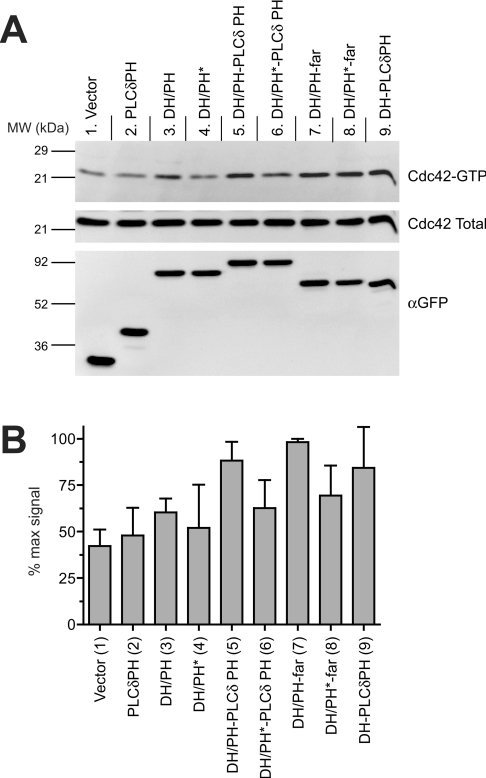
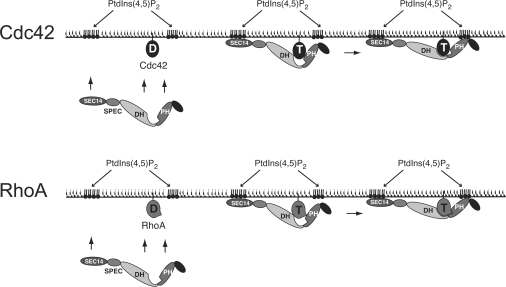
Dbl family GEFs (guanine nucleotide-exchange factors) for the Rho GTPases almost invariably contain a PH (pleckstrin homology) domain adjacent to their DH (Dbl homology) domain. The DH domain is responsible for GEF activity, and the PH domain plays a regulatory role that remains poorly understood. We demonstrated previously that Dbl family PH domains bind phosphoinositides with low affinity and cannot function as independent membrane targeting modules. In the present study, we show that dimerization of a Dbs (Dbl's big sister) DH/PH domain fragment is sufficient to drive it to the plasma membrane through a mechanism involving PH domain-phosphoinositide interactions. Thus, the Dbs PH domain could play a significant role in membrane targeting if it co-operates with other domains in the protein. We also show that mutations that prevent phosphoinositide binding by the Dbs PH domain significantly impair cellular GEF activity even in chimaeric proteins that are robustly membrane targeted by farnesylation or by the PH domain of phospholipase C-delta1. This finding argues that the Dbs PH domain plays a regulatory role that is independent of its ability to aid membrane targeting. Thus, we suggest that the PH domain plays dual roles, contributing independently to membrane localization of Dbs (as part of a multi-domain interaction) and allosteric regulation of the DH domain.
Figures
References
-
- Whitehead I. P., Campbell S., Rossman K. L., Der C. J. Dbl family proteins. Biochim. Biophys. Acta. 1997;1332:F1–F23. - PubMed
-
- Hall A. Rho GTPases and the actin cytoskeleton. Science. 1998;279:509–514. - PubMed
-
- Hart M. J., Eva A., Zangrilli D., Aaronson S. A., Evans T., Cerione R. A., Zheng Y. Cellular transformation and guanine nucleotide exchange activity are catalysed by a common domain on the dbl oncogene product. J. Biol. Chem. 1994;269:62–65. - PubMed
-
- Snyder J. T., Worthylake D. K., Rossman K. L., Betts L., Pruitt W. M., Siderovski D. P., Der C. J., Sondek J. Structural basis for the selective activation of Rho GTPases by Dbl exchange factors. Nat. Struct. Biol. 2002;9:468–475. - PubMed
-
- Rossman K. L., Sondek J. Larger than Dbl: new structural insights into RhoA activation. Trends Biochem. Sci. 2005;30:163–165. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases