

The origin of replicators and reproducers
- PMID: 17008217
- PMCID: PMC1664675
- DOI: 10.1098/rstb.2006.1912
The origin of replicators and reproducers
Abstract
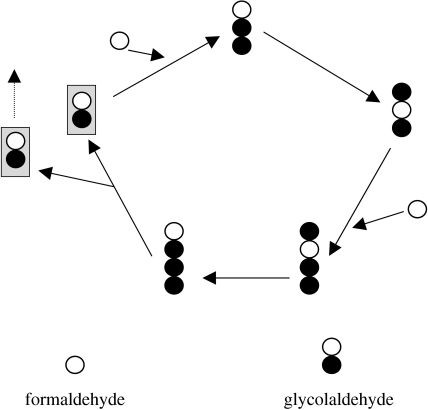
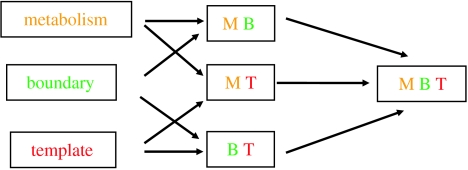
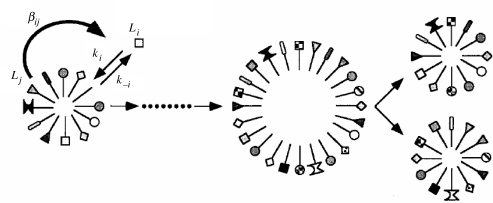
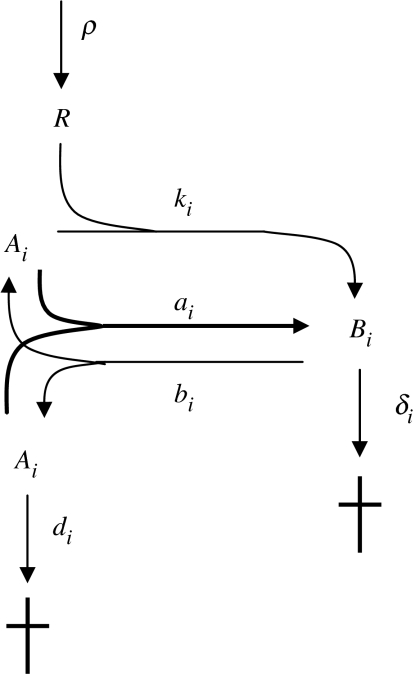
Replicators are fundamental to the origin of life and evolvability. Their survival depends on the accuracy of replication and the efficiency of growth relative to spontaneous decay. Infrabiological systems are built of two coupled autocatalytic systems, in contrast to minimal living systems that must comprise at least a metabolic subsystem, a hereditary subsystem and a boundary, serving respective functions. Some scenarios prefer to unite all these functions into one primordial system, as illustrated in the lipid world scenario, which is considered as a didactic example in detail. Experimentally produced chemical replicators grow parabolically owing to product inhibition. A selection consequence is survival of everybody. The chromatographized replicator model predicts that such replicators spreading on surfaces can be selected for higher replication rate because double strands are washed away slower than single strands from the surface. Analysis of real ribozymes suggests that the error threshold of replication is less severe by about one order of magnitude than thought previously. Surface-bound dynamics is predicted to play a crucial role also for exponential replicators: unlinked genes belonging to the same genome do not displace each other by competition, and efficient and accurate replicases can spread. The most efficient form of such useful population structure is encapsulation by reproducing vesicles. The stochastic corrector model shows how such a bag of genes can survive, and what the role of chromosome formation and intragenic recombination could be. Prebiotic and early evolution cannot be understood without the models of dynamics.
Figures
Similar articles
-
Survival of replicators with parabolic growth tendency and exponential decay.J Theor Biol. 2001 Sep 7;212(1):99-105. doi: 10.1006/jtbi.2001.2360. J Theor Biol. 2001. PMID: 11527448
-
Metabolically Coupled Replicator Systems: Overview of an RNA-world model concept of prebiotic evolution on mineral surfaces.J Theor Biol. 2015 Sep 21;381:39-54. doi: 10.1016/j.jtbi.2015.06.002. Epub 2015 Jun 15. J Theor Biol. 2015. PMID: 26087284 Review.
-
Coevolution of reproducers and replicators at the origin of life and the conditions for the origin of genomes.Proc Natl Acad Sci U S A. 2023 Apr 4;120(14):e2301522120. doi: 10.1073/pnas.2301522120. Epub 2023 Mar 30. Proc Natl Acad Sci U S A. 2023. PMID: 36996101 Free PMC article.
-
Lack of evolvability in self-sustaining autocatalytic networks constraints metabolism-first scenarios for the origin of life.Proc Natl Acad Sci U S A. 2010 Jan 26;107(4):1470-5. doi: 10.1073/pnas.0912628107. Epub 2010 Jan 4. Proc Natl Acad Sci U S A. 2010. PMID: 20080693 Free PMC article.
-
Ecology and Evolution in the RNA World Dynamics and Stability of Prebiotic Replicator Systems.Life (Basel). 2017 Nov 27;7(4):48. doi: 10.3390/life7040048. Life (Basel). 2017. PMID: 29186916 Free PMC article. Review.
Cited by
-
Hypercycle.PLoS Comput Biol. 2016 Apr 7;12(4):e1004853. doi: 10.1371/journal.pcbi.1004853. eCollection 2016 Apr. PLoS Comput Biol. 2016. PMID: 27054759 Free PMC article. No abstract available.
-
Selectionist and evolutionary approaches to brain function: a critical appraisal.Front Comput Neurosci. 2012 Apr 26;6:24. doi: 10.3389/fncom.2012.00024. eCollection 2012. Front Comput Neurosci. 2012. PMID: 22557963 Free PMC article.
-
A new replicator: a theoretical framework for analysing replication.BMC Biol. 2010 Mar 10;8:21. doi: 10.1186/1741-7007-8-21. BMC Biol. 2010. PMID: 20219099 Free PMC article.
-
Synthetic Mutualism and the Intervention Dilemma.Life (Basel). 2019 Jan 28;9(1):15. doi: 10.3390/life9010015. Life (Basel). 2019. PMID: 30696090 Free PMC article. Review.
-
The essence of life.Biol Direct. 2016 Sep 26;11(1):49. doi: 10.1186/s13062-016-0150-5. Biol Direct. 2016. PMID: 27671203 Free PMC article.
References
-
- Bedau M.A, McCaskill J.S, Packard N.H, Rasmussen S, Adami C, Green D.G, Ikegami T, Kaneko K, Ray T.S. Open problems in artificial life. Artif. Life. 2000;6:363–376. doi:10.1162/106454600300103683 - DOI - PubMed
-
- Boerlijst M.C, Hogeweg P. Spiral wave structure in pre-biotic evolution—hypercycles stable against parasites. Physica. 1991;D48:17–28.
-
- Czárán T, Szathmáry E. Coexistence of replicators in prebiotic evolution. In: Dieckmann U, Law R, Metz J.A.J, editors. The geometry of ecological interactions: simplifying spatial complexity. IIASA and Cambridge University Press; Wien, Austria: 2000. pp. 116–134.
-
- Dawkins R. The selfish gene. Oxford University Press; Oxford, UK: 1976.
-
- Eigen M. Self-organization of matter and the evolution of biological macromolecules. Naturwissenschaften. 1971;58:465–523. doi:10.1007/BF00623322 - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
