

The origami of thioredoxin-like folds
- PMID: 17008712
- PMCID: PMC2242382
- DOI: 10.1110/ps.062268106
The origami of thioredoxin-like folds
Abstract
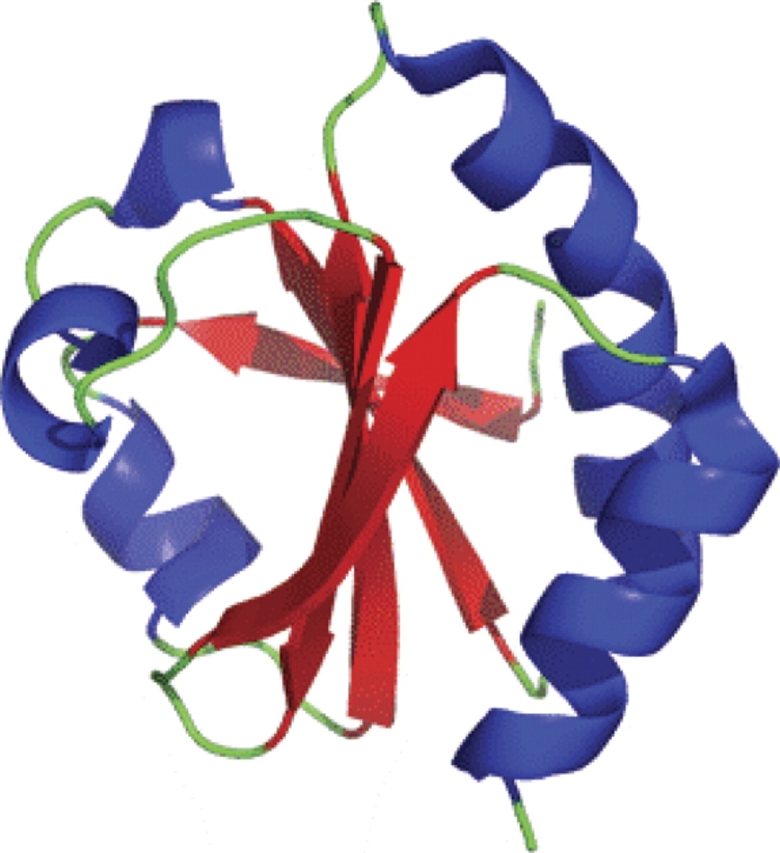
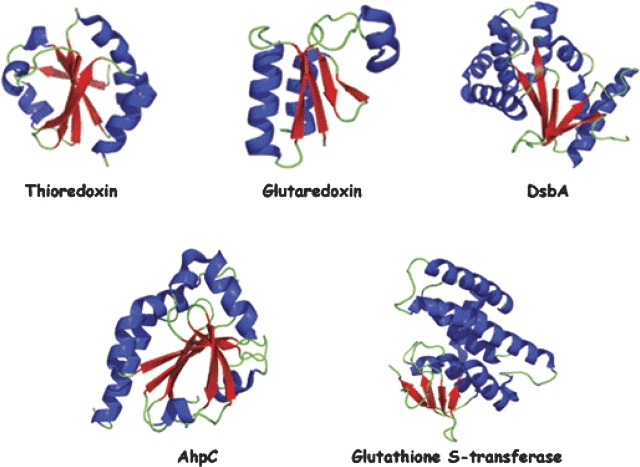
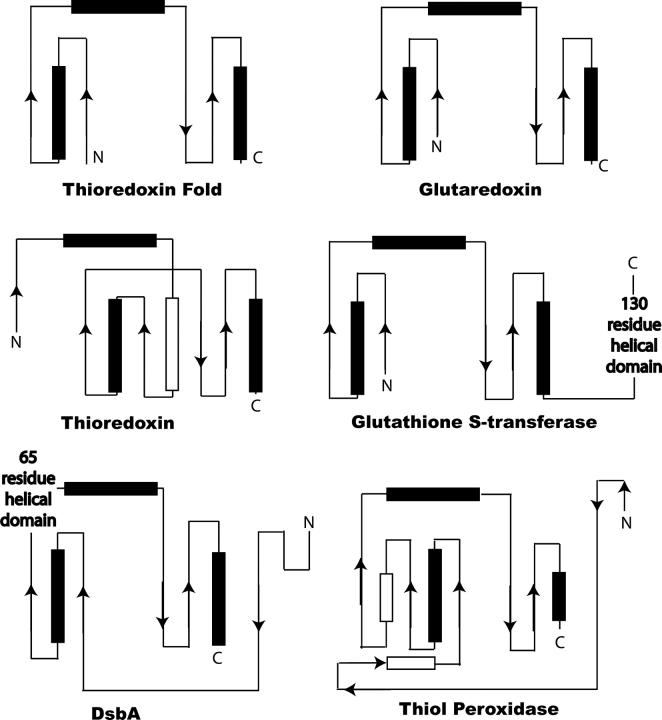
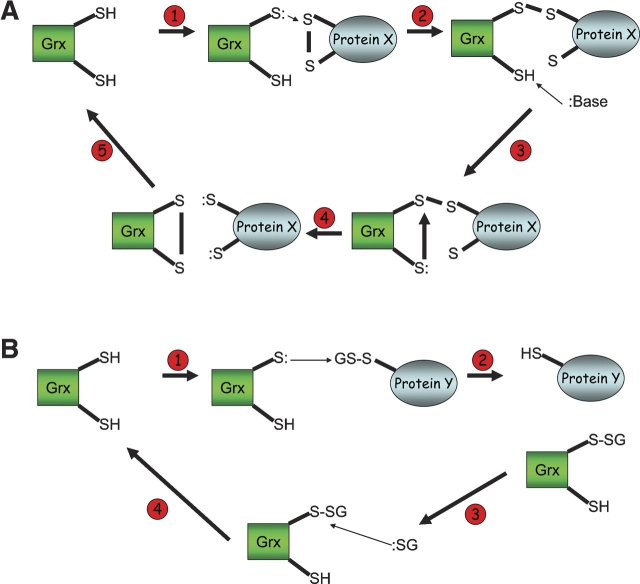
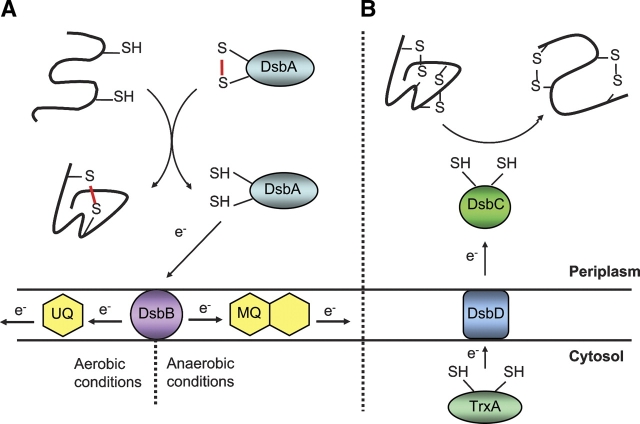
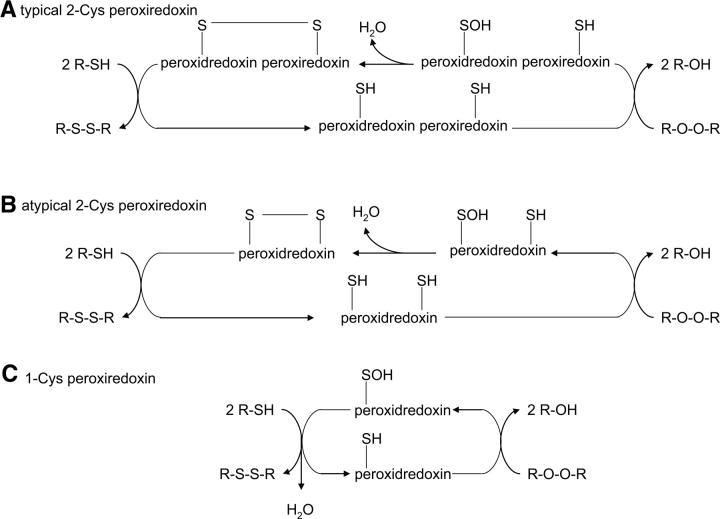
Origami is the Japanese art of folding a piece of paper into complex shapes and forms. Much like origami of paper, Nature has used conserved protein folds to engineer proteins for a particular task. An example of a protein family, which has been used by Nature numerous times, is the thioredoxin superfamily. Proteins in the thioredoxin superfamily are all structured with a beta-sheet core surrounded with alpha-helices, and most contain a canonical CXXC motif. The remarkable feature of these proteins is that the link between them is the fold; however, their reactivity is different for each member due to small variations in this general fold as well as their active site. This review attempts to unravel the minute differences within this protein family, and it also demonstrates the ingenuity of Nature to use a conserved fold to generate a diverse collection of proteins to perform a number of different biochemical tasks.
Figures

Similar articles
-
Structural and functional characterization of a thioredoxin-like protein (Mt0807) from Methanobacterium thermoautotrophicum.Biochemistry. 2003 Jul 8;42(26):8001-10. doi: 10.1021/bi030021g. Biochemistry. 2003. PMID: 12834352
-
Structural classification of thioredoxin-like fold proteins.Proteins. 2005 Feb 1;58(2):376-88. doi: 10.1002/prot.20329. Proteins. 2005. PMID: 15558583
-
A conserved cis-proline precludes metal binding by the active site thiolates in members of the thioredoxin family of proteins.Biochemistry. 2007 Jun 12;46(23):6903-10. doi: 10.1021/bi700152b. Epub 2007 May 16. Biochemistry. 2007. PMID: 17503777
-
Recruitment of thioredoxin-like domains into prostaglandin synthases.Biochem Biophys Res Commun. 2008 May 2;369(2):281-6. doi: 10.1016/j.bbrc.2008.02.088. Epub 2008 Feb 26. Biochem Biophys Res Commun. 2008. PMID: 18307977 Review.
-
Similarities and differences in the thioredoxin superfamily.Prog Biophys Mol Biol. 2006 Jul;91(3):229-48. doi: 10.1016/j.pbiomolbio.2005.06.012. Epub 2005 Jul 26. Prog Biophys Mol Biol. 2006. PMID: 16098567 Review.
Cited by
-
Fort CnoX: Protecting Bacterial Proteins From Misfolding and Oxidative Damage.Front Mol Biosci. 2021 May 4;8:681932. doi: 10.3389/fmolb.2021.681932. eCollection 2021. Front Mol Biosci. 2021. PMID: 34017858 Free PMC article. Review.
-
Unraveling the roles of aromatic cluster side-chain interactions on the structural stability and functional significance of psychrophilic Sphingomonas sp. glutaredoxin 3.PLoS One. 2023 Aug 31;18(8):e0290686. doi: 10.1371/journal.pone.0290686. eCollection 2023. PLoS One. 2023. PMID: 37651358 Free PMC article.
-
A Disulfide Oxidoreductase (CHU_1165) Is Essential for Cellulose Degradation by Affecting Outer Membrane Proteins in Cytophaga hutchinsonii.Appl Environ Microbiol. 2020 Apr 1;86(8):e02789-19. doi: 10.1128/AEM.02789-19. Print 2020 Apr 1. Appl Environ Microbiol. 2020. PMID: 32033954 Free PMC article.
-
Biosynthesis of rhizocticins, antifungal phosphonate oligopeptides produced by Bacillus subtilis ATCC6633.Chem Biol. 2010 Jan 29;17(1):28-37. doi: 10.1016/j.chembiol.2009.11.017. Chem Biol. 2010. PMID: 20142038 Free PMC article.
-
Topological variation in the evolution of new reactions in functionally diverse enzyme superfamilies.Curr Opin Struct Biol. 2011 Jun;21(3):391-7. doi: 10.1016/j.sbi.2011.03.007. Epub 2011 Apr 1. Curr Opin Struct Biol. 2011. PMID: 21458983 Free PMC article. Review.
References
-
- Andersen, C.L., Matthey-Dupraz, A., Missiakas, D., Raina, S. 1997. A new Escherichia coli gene, dsbG, encodes a periplasmic protein involved in disulphide bond formation, required for recycling DsbA/DsbB and DsbC redox proteins. Mol. Microbiol. 26: 121–132. - PubMed
-
- Aslund, F., Nordstrand, K., Berndt, K.D., Nikkola, M., Bergman, T., Ponstingl, H., Jornvall, H., Otting, G., Holmgren, A. 1996. Glutaredoxin-3 from Escherichia coli: Amino acid sequence, 1H and 15N NMR assignments, and structural analysis. J. Biol. Chem. 271: 6736–6745. - PubMed
-
- Aslund, F., Berndt, K.D., Holmgren, A. 1997. Redox potentials of glutaredoxins and other thiol-disulfide oxidoreductases of the thioredoxin superfamily determined by direct protein–protein redox equilibria. J. Biol. Chem. 272: 30780–30786. - PubMed
-
- Bader, M., Muse, W., Ballou, D.P., Gassner, C., Bardwell, J.C. 1999. Oxidative protein folding is driven by the electron transport system. Cell 98: 217–227. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
