

Combinatorial Domain Hunting: An effective approach for the identification of soluble protein domains adaptable to high-throughput applications
- PMID: 17008718
- PMCID: PMC2242398
- DOI: 10.1110/ps.062082606
Combinatorial Domain Hunting: An effective approach for the identification of soluble protein domains adaptable to high-throughput applications
Abstract
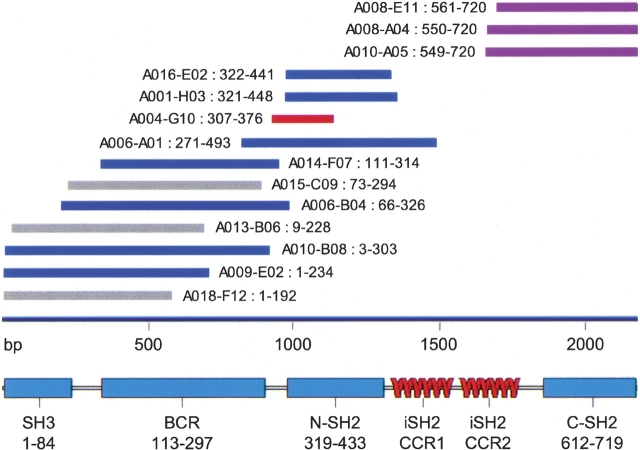
Exploitation of potential new targets for drug and vaccine development has an absolute requirement for multimilligram quantities of soluble protein. While recombinant expression of full-length proteins is frequently problematic, high-yield soluble expression of functional subconstructs is an effective alternative, so long as appropriate termini can be identified. Bioinformatics localizes domains, but doesn't predict boundaries with sufficient accuracy, so that subconstructs are typically found by trial and error. Combinatorial Domain Hunting (CDH) is a technology for discovering soluble, highly expressed constructs of target proteins. CDH combines unbiased, finely sampled gene-fragment libraries, with a screening protocol that provides "holistic" readout of solubility and yield for thousands of protein fragments. CDH is free of the "passenger solubilization" and out-of-frame translational start artifacts of fusion-protein systems, and hits are ready for scale-up expression. As a proof of principle, we applied CDH to p85alpha, successfully identifying soluble and highly expressed constructs encapsulating all the known globular domains, and immediately suitable for downstream applications.
Figures

Similar articles
-
DNA fragmentation based combinatorial approaches to soluble protein expression Part II: library expression, screening and scale-up.Drug Discov Today. 2007 Nov;12(21-22):939-47. doi: 10.1016/j.drudis.2007.08.016. Epub 2007 Oct 22. Drug Discov Today. 2007. PMID: 17993412
-
DNA fragmentation-based combinatorial approaches to soluble protein expression Part I. Generating DNA fragment libraries.Drug Discov Today. 2007 Nov;12(21-22):931-8. doi: 10.1016/j.drudis.2007.08.012. Epub 2007 Oct 23. Drug Discov Today. 2007. PMID: 17993411 Review.
-
ORF-selector ESPRIT: a second generation library screen for soluble protein expression employing precise open reading frame selection.J Struct Biol. 2011 Aug;175(2):189-97. doi: 10.1016/j.jsb.2011.04.004. Epub 2011 Apr 16. J Struct Biol. 2011. PMID: 21515383
-
ESPRIT: an automated, library-based method for mapping and soluble expression of protein domains from challenging targets.J Struct Biol. 2010 Oct;172(1):66-74. doi: 10.1016/j.jsb.2010.02.021. Epub 2010 Mar 4. J Struct Biol. 2010. PMID: 20206698 Review.
-
Combinatorial Domain Hunting: solving problems in protein expression.Drug Discov Today. 2010 Jun;15(11-12):461-7. doi: 10.1016/j.drudis.2010.03.010. Epub 2010 Mar 23. Drug Discov Today. 2010. PMID: 20338258 Review.
Cited by
-
Library methods for structural biology of challenging proteins and their complexes.Curr Opin Struct Biol. 2013 Jun;23(3):403-8. doi: 10.1016/j.sbi.2013.03.004. Epub 2013 Apr 17. Curr Opin Struct Biol. 2013. PMID: 23602357 Free PMC article. Review.
-
Small-scale, semi-automated purification of eukaryotic proteins for structure determination.J Struct Funct Genomics. 2007 Dec;8(4):153-66. doi: 10.1007/s10969-007-9032-5. Epub 2007 Nov 6. J Struct Funct Genomics. 2007. PMID: 17985212 Free PMC article.
-
Protein stability: a crystallographer's perspective.Acta Crystallogr F Struct Biol Commun. 2016 Feb;72(Pt 2):72-95. doi: 10.1107/S2053230X15024619. Epub 2016 Jan 26. Acta Crystallogr F Struct Biol Commun. 2016. PMID: 26841758 Free PMC article. Review.
-
Experimental mapping of soluble protein domains using a hierarchical approach.Nucleic Acids Res. 2011 Oct;39(18):e125. doi: 10.1093/nar/gkr548. Epub 2011 Jul 19. Nucleic Acids Res. 2011. PMID: 21771856 Free PMC article.
-
A high-throughput immobilized bead screen for stable proteins and multi-protein complexes.Protein Eng Des Sel. 2011 Jul;24(7):565-78. doi: 10.1093/protein/gzr021. Epub 2011 Jun 3. Protein Eng Des Sel. 2011. PMID: 21642284 Free PMC article.
References
-
- Blundell, T.L., Jhoti, H., Abell, C. 2002. High-throughput crystallography for lead discovery in drug design. Nat. Rev. Drug Discov. 1: 45–54. - PubMed
-
- Booker, G.W., Breeze, A.L., Downing, A.K., Panayotou, G., Gout, I., Waterfield, M.D., Campbell, I.D. 1992. Structure of an SH2 domain of the p85 α subunit of phosphatidylinositol-3-OH kinase. Nature 358: 684–687. - PubMed
-
- Booker, G.W., Gout, I., Downing, A.K., Driscoll, P.C., Boyd, J., Waterfield, M.D., Campbell, I.D. 1993. Solution structure and ligand-binding site of the SH3 domain of the p85 α subunit of phosphatidylinositol 3-kinase. Cell 73: 813–822. - PubMed
-
- Cabantous, S., Pedelacq, J.D., Mark, B.L., Naranjo, C., Terwilliger, T.C., Waldo, G.S. 2005a. Recent advances in GFP folding reporter and split-GFP solubility reporter technologies. Application to improving the folding and solubility of recalcitrant proteins from Mycobacterium tuberculosis . J. Struct. Funct. Genomics 6: 113–119. - PubMed
-
- Cabantous, S., Terwilliger, T.C., Waldo, G.S. 2005b. Protein tagging and detection with engineered self-assembling fragments of green fluorescent protein. Nat. Biotechnol. 23: 102–107. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
