

Engineered zinc-binding sites confirm proximity and orientation of transmembrane helices I and III in the human serotonin transporter
- PMID: 17008722
- PMCID: PMC2242390
- DOI: 10.1110/ps.062386106
Engineered zinc-binding sites confirm proximity and orientation of transmembrane helices I and III in the human serotonin transporter
Abstract
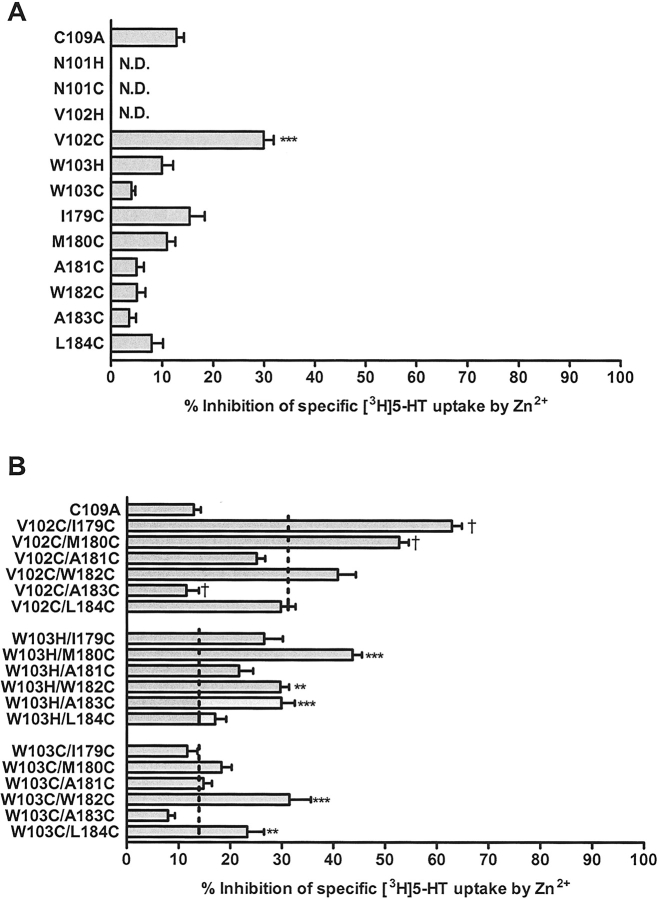
The human serotonin transporter (hSERT) regulates neurotransmission by removing released serotonin (5-HT) from the synapse. Previous studies identified residues in SERT transmembrane helices (TMHs) I and III as interaction sites for substrates and antagonists. Despite an abundance of data supporting a 12-TMH topology, the arrangement of the TMHs in SERT and other biogenic amine transporters remains undetermined. A high-resolution structure of a bacterial leucine transporter that demonstrates homology with SERT has been reported, thus providing the basis for the development of a SERT model. Zn2+-binding sites have been utilized in transporters and receptors to define experimentally TMH proximity. Focusing on residues near the extracellular ends of hSERT TMHs I and III, we engineered potential Zn2+-binding sites between V102 or W103 (TMH I) and I179-L184 (TMH III). Residues were mutated to either histidine or cysteine. TMH I/III double mutants were constructed from functional TMH I mutants, and Zn2+ sensitivity was assessed. Dose-response assays suggest an approximately twofold increase in sensitivity to Zn2+ inhibition at the hSERT V102C/M180C and approximately fourfold at the V102C/I179C mutant compared to the hSERT V102C single mutant. We propose that the increased sensitivity to Zn2+ confirms the proximity and the orientation of TMHs I and III in the membrane. Homology modeling of the proposed Zn2+-binding sites using the coordinates of the Aquifex aeolicus leucine transporter structure provided a structural basis for interpreting the results and developing conclusions.
Figures







Similar articles
-
Comparative molecular field analysis using selectivity fields reveals residues in the third transmembrane helix of the serotonin transporter associated with substrate and antagonist recognition.J Pharmacol Exp Ther. 2008 Jun;325(3):791-800. doi: 10.1124/jpet.108.136200. Epub 2008 Mar 19. J Pharmacol Exp Ther. 2008. PMID: 18354055 Free PMC article.
-
Conformational flexibility of transmembrane helix VII of the human serotonin transporter impacts ion dependence and transport.Biochem Pharmacol. 2010 Nov 1;80(9):1418-26. doi: 10.1016/j.bcp.2010.07.005. Epub 2010 Jul 15. Biochem Pharmacol. 2010. PMID: 20637736 Free PMC article.
-
Binding of serotonin to the human serotonin transporter. Molecular modeling and experimental validation.J Am Chem Soc. 2008 Mar 26;130(12):3853-65. doi: 10.1021/ja076403h. Epub 2008 Mar 4. J Am Chem Soc. 2008. PMID: 18314975
-
Y95 and E444 interaction required for high-affinity S-citalopram binding in the human serotonin transporter.ACS Chem Neurosci. 2011 Feb 16;2(2):75-81. doi: 10.1021/cn100066p. Epub 2010 Oct 27. ACS Chem Neurosci. 2011. PMID: 22778858 Free PMC article. Review.
-
Structure/function relationships in serotonin transporter: new insights from the structure of a bacterial transporter.Handb Exp Pharmacol. 2006;(175):59-73. doi: 10.1007/3-540-29784-7_3. Handb Exp Pharmacol. 2006. PMID: 16722230 Review.
Cited by
-
1-Methylpyridinium-4-(4-phenylmethanethiosulfonate) iodide, MTS-MPP+, a novel scanning cysteine accessibility method (SCAM) reagent for monoamine transporter studies.Bioorg Med Chem. 2007 Jan 1;15(1):305-11. doi: 10.1016/j.bmc.2006.09.058. Epub 2006 Sep 29. Bioorg Med Chem. 2007. PMID: 17064910 Free PMC article.
-
Zinc and zinc chelators modify taurine transport in rat retinal cells.Neurochem Res. 2014 Nov;39(11):2234-9. doi: 10.1007/s11064-014-1425-9. Epub 2014 Sep 3. Neurochem Res. 2014. PMID: 25182776
-
Comparative molecular field analysis using selectivity fields reveals residues in the third transmembrane helix of the serotonin transporter associated with substrate and antagonist recognition.J Pharmacol Exp Ther. 2008 Jun;325(3):791-800. doi: 10.1124/jpet.108.136200. Epub 2008 Mar 19. J Pharmacol Exp Ther. 2008. PMID: 18354055 Free PMC article.
-
Steric hindrance mutagenesis in the conserved extracellular vestibule impedes allosteric binding of antidepressants to the serotonin transporter.J Biol Chem. 2012 Nov 16;287(47):39316-26. doi: 10.1074/jbc.M112.371765. Epub 2012 Sep 24. J Biol Chem. 2012. PMID: 23007398 Free PMC article.
-
The binding sites for cocaine and dopamine in the dopamine transporter overlap.Nat Neurosci. 2008 Jul;11(7):780-9. doi: 10.1038/nn.2146. Epub 2008 Jun 22. Nat Neurosci. 2008. PMID: 18568020 Free PMC article.
References
-
- Adkins, E.M., Barker, E.L., Blakely, R.D. 2001. Interactions of tryptamine derivatives with serotonin transporter species variants implicate transmembrane domain I in substrate recognition. Mol. Pharmacol. 59: 514–523. - PubMed
-
- Barker, E.L., Perlman, M.A., Adkins, E.M., Houlihan, W.J., Pristupa, Z.B., Niznik, H.B., Blakely, R.D. 1998. High affinity recognition of serotonin transporter antagonists defined by species-scanning mutagenesis. An aromatic residue in transmembrane domain I dictates species-selective recognition of citalopram and mazindol. J. Biol. Chem. 273: 19459–19468. - PubMed
-
- Bismuth, Y., Kavanaugh, M.P., Kanner, B.I. 1997. Tyrosine 140 of the γ-aminobutyric acid transporter GAT-1 plays a critical role in neurotransmitter recognition. J. Biol. Chem. 272: 16096–16102. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
