

Role of the transcription factor E2F1 in CXCR4-mediated neurotoxicity and HIV neuropathology
- PMID: 17011204
- PMCID: PMC1762194
- DOI: 10.1016/j.nbd.2006.08.004
Role of the transcription factor E2F1 in CXCR4-mediated neurotoxicity and HIV neuropathology
Abstract
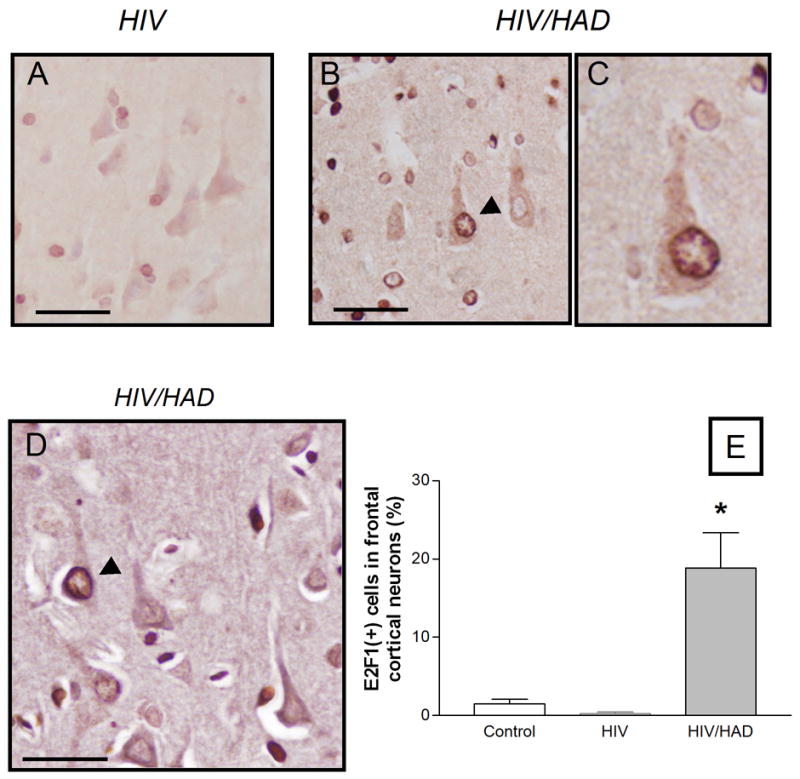
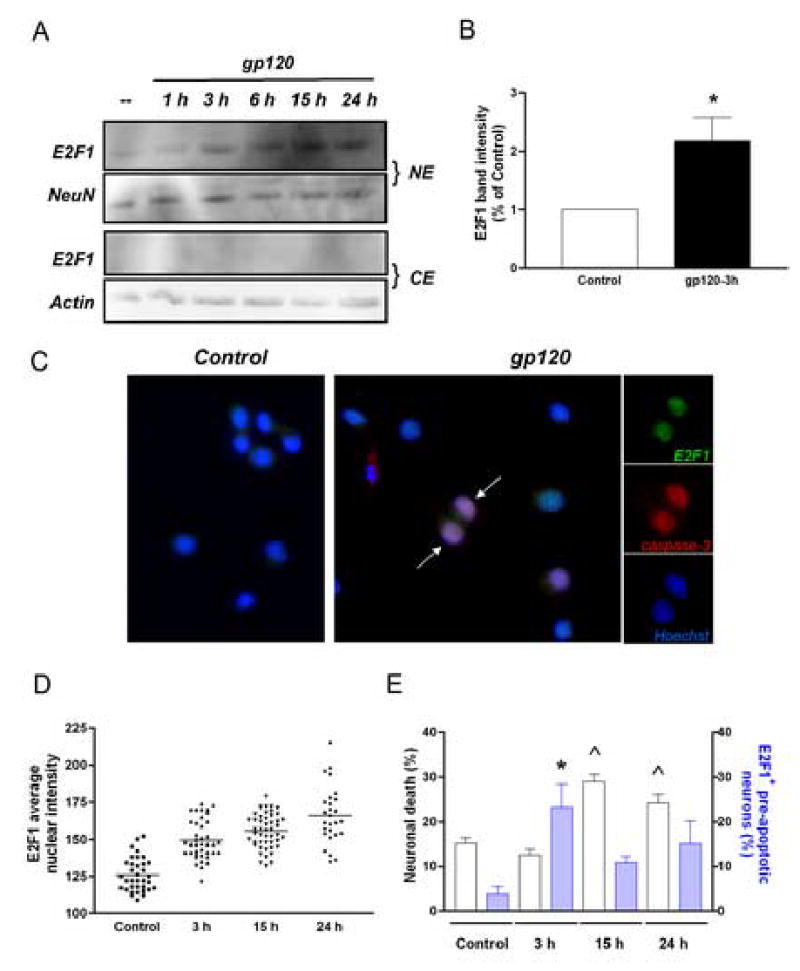
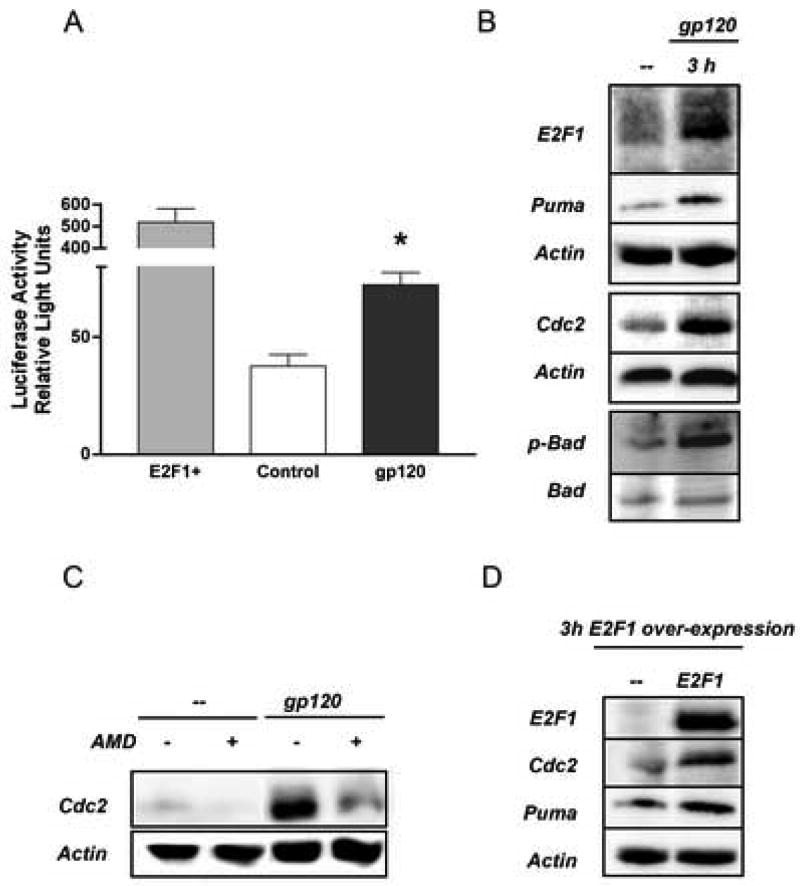
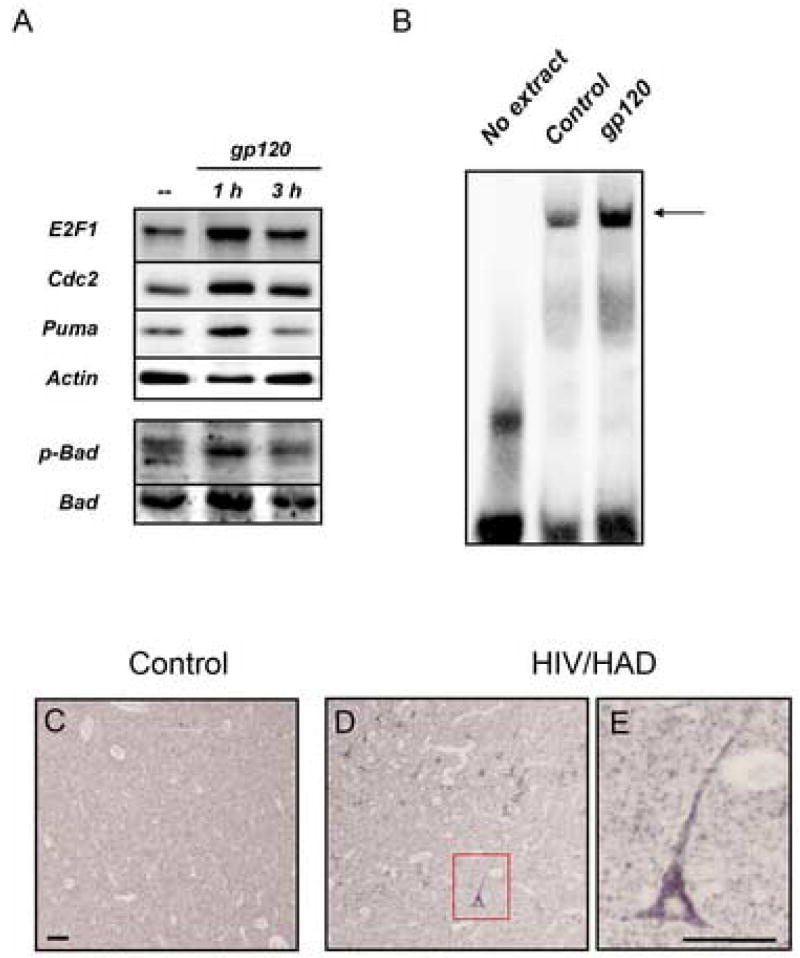
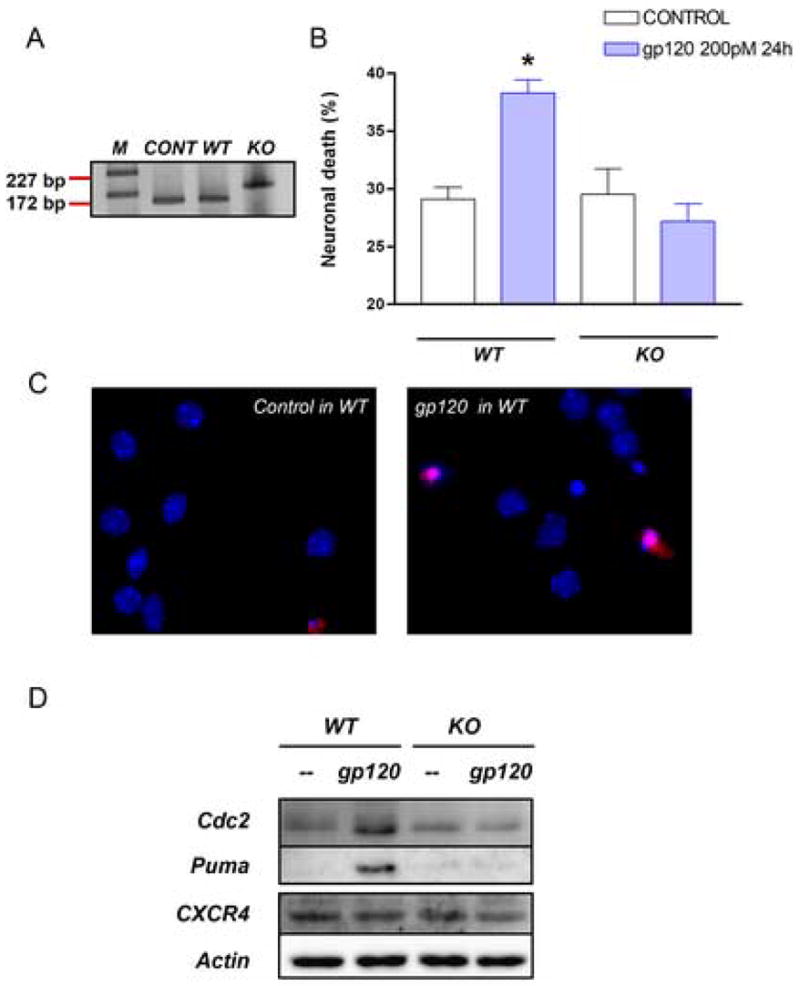
This study sought to determine the role of the transcription factor E2F1 in CXCR4-mediated neurotoxicity and HIV neuropathology. We studied the effect of the HIV envelope protein gp120 on the expression of E2F1-dependent apoptotic proteins in human and rodent neurons and examined the expression pattern of E2F1 in the brain of HIV-infected individuals. Our findings suggest that in cultured neurons gp120 increased E2F1 levels in the nucleus, stimulated its transcriptional activity and enhanced the expression of the E2F1 target proteins Cdc2 and Puma. Studies with neuronal cultures from E2F1 deficient mice demonstrated that the transcription factor is required for gp120-induced neurotoxicity and up-regulation of Cdc2 and Puma. Levels of E2F1 protein were greater in the nucleus of neurons in brains of HIV-infected patients exhibiting dementia when compared to HIV-negative subjects or HIV-positive neurologically normal patients. Overall, these studies indicate that E2F1 is primarily involved in CXCR4-mediated neurotoxicity and HIV neuropathogenesis.
Figures
References
-
- Adle-Biassette H, Levy Y, Colombel M, Poron F, Natchev S, Keohane C, Gray F. Neuronal apoptosis in HIV infection in adults. Neuropathol Appl Neurobiol. 1995;21:218–227. - PubMed
-
- Cartier L, Hartley O, Dubois-Dauphin M, Krause KH. Chemokine receptors in the central nervous system: role in brain inflammation and neurodegenerative diseases. Brain Res Brain Res Rev. 2005;48:16–42. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- MH59724/MH/NIMH NIH HHS/United States
- R24 MH059724/MH/NIMH NIH HHS/United States
- R01 DA015014/DA/NIDA NIH HHS/United States
- R24 MH059745/MH/NIMH NIH HHS/United States
- R24 NS038841/NS/NINDS NIH HHS/United States
- NS 41561/NS/NINDS NIH HHS/United States
- R01 NS041561/NS/NINDS NIH HHS/United States
- DA 19808/DA/NIDA NIH HHS/United States
- NS45491/NS/NINDS NIH HHS/United States
- NS38841/NS/NINDS NIH HHS/United States
- R37 DA015014/DA/NIDA NIH HHS/United States
- DA15014/DA/NIDA NIH HHS/United States
- MH59745-06/MH/NIMH NIH HHS/United States
- R24 NS045491/NS/NINDS NIH HHS/United States
- R01 DA019808/DA/NIDA NIH HHS/United States
- N01 MH032002/MH/NIMH NIH HHS/United States
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
