

Agrobacterium rhizogenes GALLS protein contains domains for ATP binding, nuclear localization, and type IV secretion
- PMID: 17012398
- PMCID: PMC1698208
- DOI: 10.1128/JB.00747-06
Agrobacterium rhizogenes GALLS protein contains domains for ATP binding, nuclear localization, and type IV secretion
Abstract
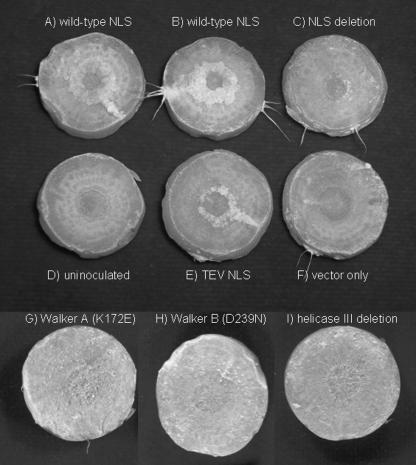
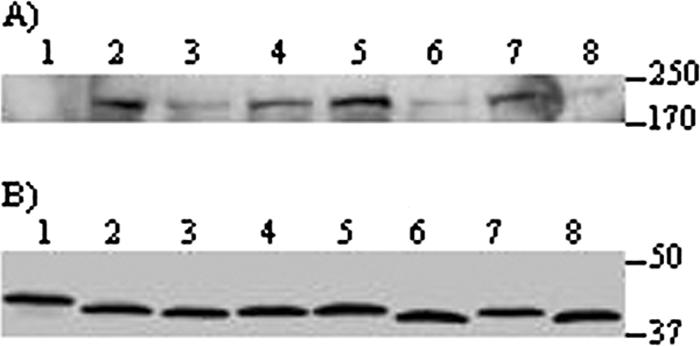
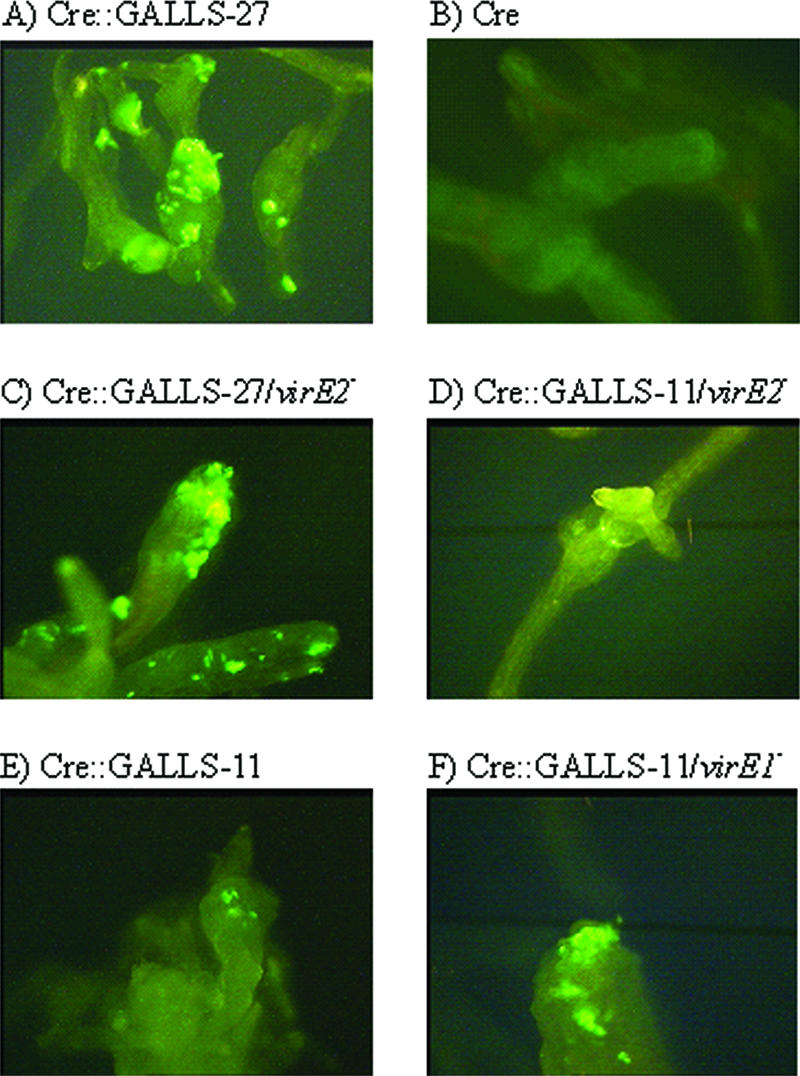
Agrobacterium tumefaciens and Agrobacterium rhizogenes are closely related plant pathogens that cause different diseases, crown gall and hairy root. Both diseases result from transfer, integration, and expression of plasmid-encoded bacterial genes located on the transferred DNA (T-DNA) in the plant genome. Bacterial virulence (Vir) proteins necessary for infection are also translocated into plant cells. Transfer of single-stranded DNA (ssDNA) and Vir proteins requires a type IV secretion system, a protein complex spanning the bacterial envelope. A. tumefaciens translocates the ssDNA-binding protein VirE2 into plant cells, where it binds single-stranded T-DNA and helps target it to the nucleus. Although some strains of A. rhizogenes lack VirE2, they are pathogenic and transfer T-DNA efficiently. Instead, these bacteria express the GALLS protein, which is essential for their virulence. The GALLS protein can complement an A. tumefaciens virE2 mutant for tumor formation, indicating that GALLS can substitute for VirE2. Unlike VirE2, GALLS contains ATP-binding and helicase motifs similar to those in TraA, a strand transferase involved in conjugation. Both GALLS and VirE2 contain nuclear localization sequences and a C-terminal type IV secretion signal. Here we show that mutations in any of these domains abolished the ability of GALLS to substitute for VirE2.
Figures

References
-
- Beijersbergen, A., A. D. Dulk-Ras, R. A. Schilperoort, and P. J. J. Hooykaas. 1992. Conjugative transfer by the virulence system of Agrobacterium tumefaciens. Science 256:1324-1327. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
