

Tocopherols play a crucial role in low-temperature adaptation and Phloem loading in Arabidopsis
- PMID: 17012603
- PMCID: PMC1626601
- DOI: 10.1105/tpc.105.039404
Tocopherols play a crucial role in low-temperature adaptation and Phloem loading in Arabidopsis
Abstract
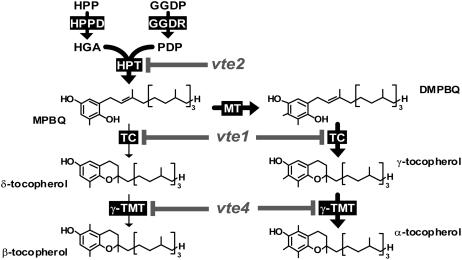
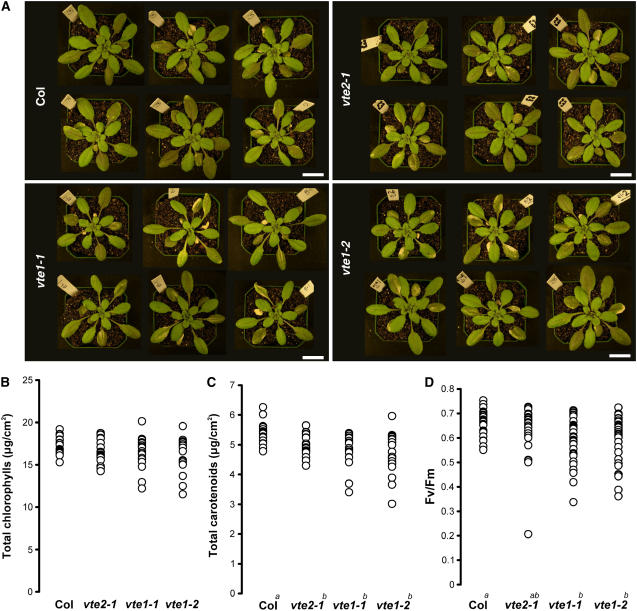
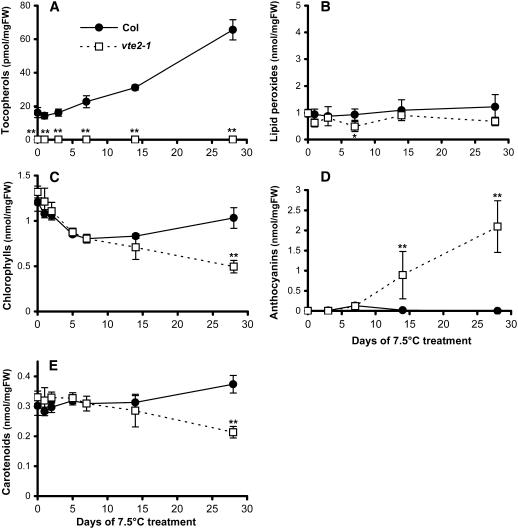
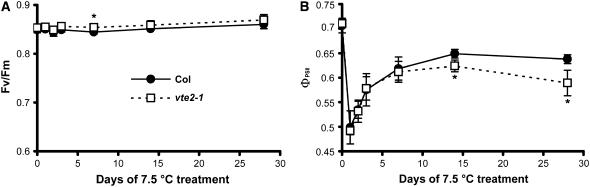
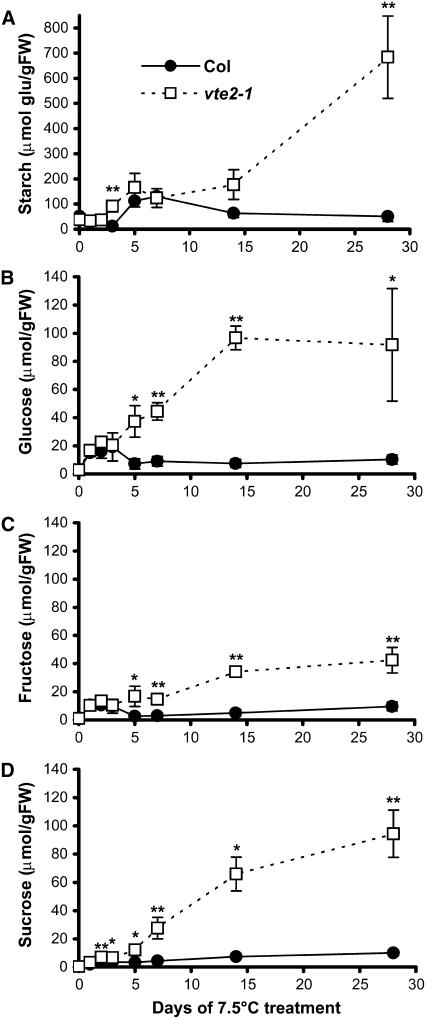
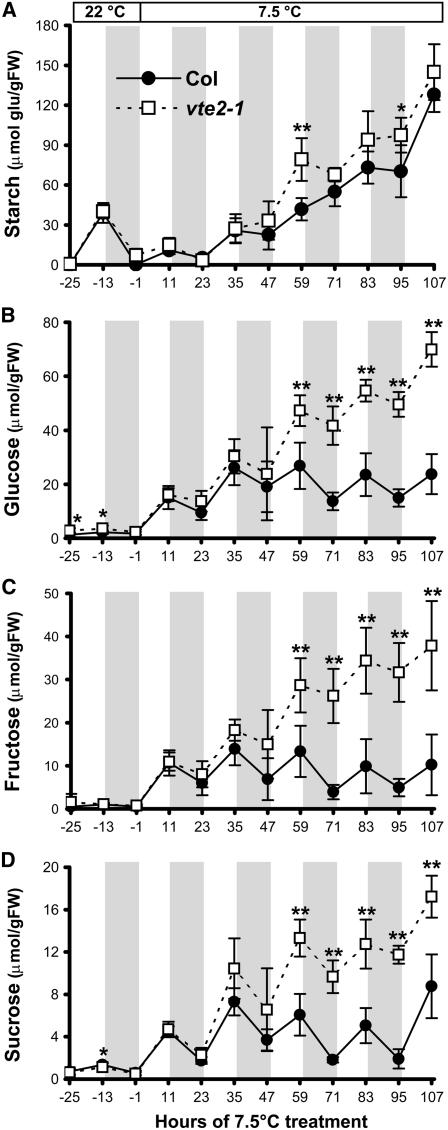
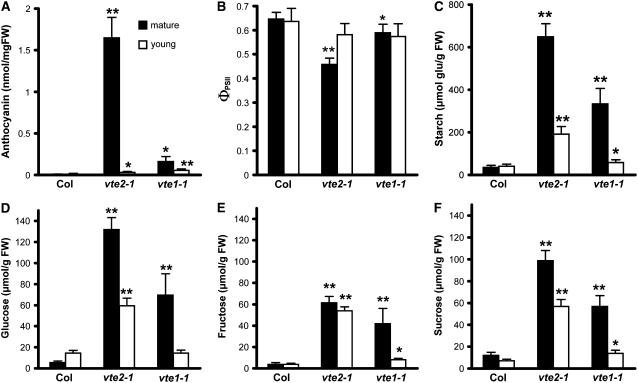
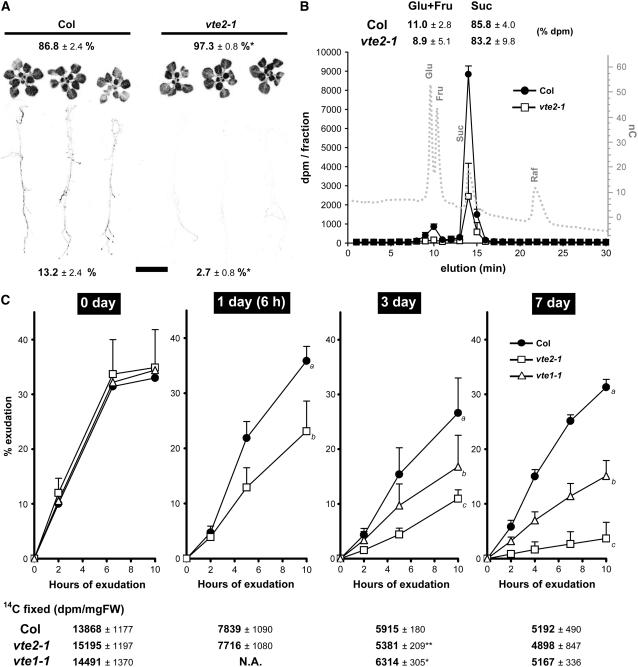
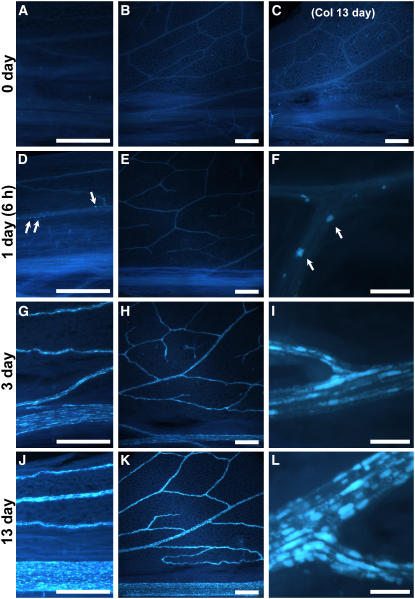
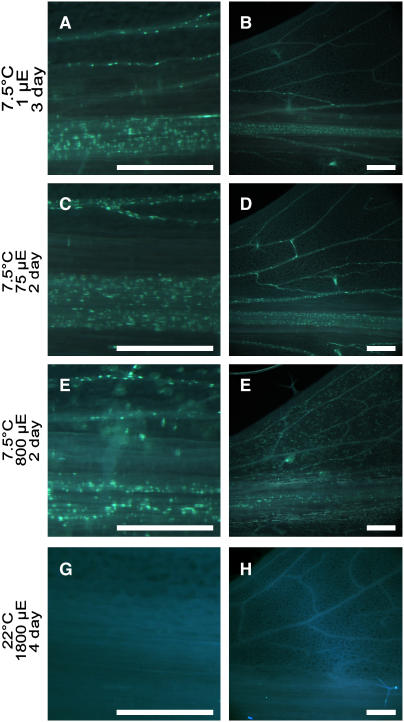
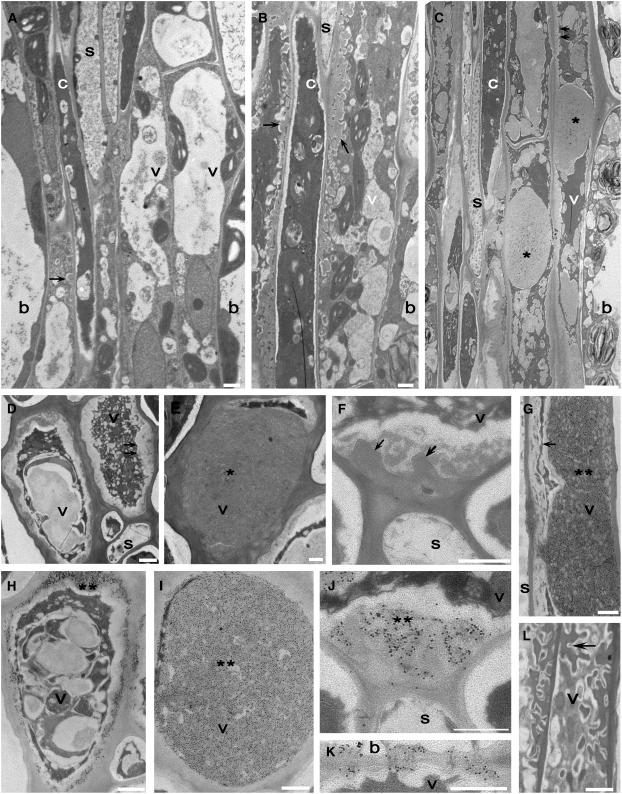
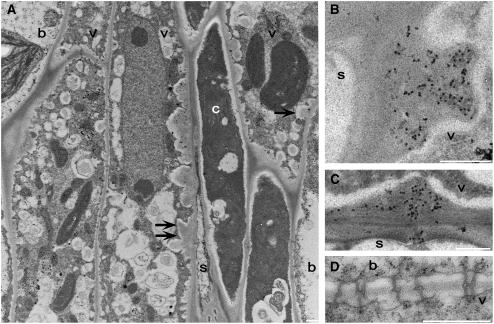
To test whether tocopherols (vitamin E) are essential in the protection against oxidative stress in plants, a series of Arabidopsis thaliana vitamin E (vte) biosynthetic mutants that accumulate different types and levels of tocopherols and pathway intermediates were analyzed under abiotic stress. Surprisingly subtle differences were observed between the tocopherol-deficient vte2 mutant and the wild type during high-light, salinity, and drought stresses. However, vte2, and to a lesser extent vte1, exhibited dramatic phenotypes under low temperature (i.e., increased anthocyanin levels and reduced growth and seed production). That these changes were independent of light level and occurred in the absence of photoinhibition or lipid peroxidation suggests that the mechanisms involved are independent of tocopherol functions in photoprotection. Compared with the wild type, vte1 and vte2 had reduced rates of photoassimilate export as early as 6 h into low-temperature treatment, increased soluble sugar levels by 60 h, and increased starch and reduced photosynthetic electron transport rate by 14 d. The rapid reduction in photoassimilate export in vte2 coincides with callose deposition exclusively in phloem parenchyma transfer cell walls adjacent to the companion cell/sieve element complex. Together, these results indicate that tocopherols have a more limited role in photoprotection than previously assumed but play crucial roles in low-temperature adaptation and phloem loading.
Figures

References
-
- Bergmuller, E., Porfirova, S., and Dormann, P. (2003). Characterization of an Arabidopsis mutant deficient in gamma-tocopherol methyltransferase. Plant Mol. Biol. 52 1181–1190. - PubMed
-
- Bjorkman, O., and Demmig, B. (1987). Photon yield of O2 evolution and chlorophyll fluorescence characteristics at 77 K among vascular plants of diverse origins. Planta 170 489–504. - PubMed
-
- Botha, C.E.J., Cross, R.H.M., van Bel, A.J.E., and Peter, C.I. (2000). Phloem loading in the sucrose-export-defective (SXD-1) mutant maize is limited by callose deposition at plasmodesmata in bundle sheath-vascular parenchyma interface. Protoplasma 214 65–72.
-
- Bradford, A., Atkinson, J., Fuller, N., and Rand, R.P. (2003). The effect of vitamin E on the structure of membrane lipid assemblies. J. Lipid Res. 44 1940–1945. - PubMed
-
- Bramley, P.M., Elmadfa, I., Kafatos, A., Kelly, F.J., Manios, Y., Roxborough, H.E., Schuch, W., Sheehy, P.J.A., and Wagner, K.H. (2000). Vitamin E. J. Sci. Food Agric. 80 913–938.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
