

The translocated Salmonella effector proteins SseF and SseG interact and are required to establish an intracellular replication niche
- PMID: 17015457
- PMCID: PMC1698088
- DOI: 10.1128/IAI.00648-06
The translocated Salmonella effector proteins SseF and SseG interact and are required to establish an intracellular replication niche
Abstract
The facultative intracellular pathogen Salmonella enterica causes a variety of diseases, including gastroenteritis and typhoid fever. Inside epithelial cells, Salmonella replicates in vacuoles, which localize in the perinuclear area in close proximity to the Golgi apparatus. Among the effector proteins translocated by the Salmonella pathogenicity island 2-encoded type III secretion system, SifA and SseG have been shown necessary but not sufficient to ensure the intracellular positioning of Salmonella vacuoles. Hence, we have investigated the involvement of other secreted effector proteins in this process. Here we show that SseF interacts functionally and physically with SseG but not SifA and is also required for the perinuclear localization of Salmonella vacuoles. The observations show that the intracellular positioning of Salmonella vacuoles is a complex phenomenon resulting from the combined action of several effector proteins.
Figures

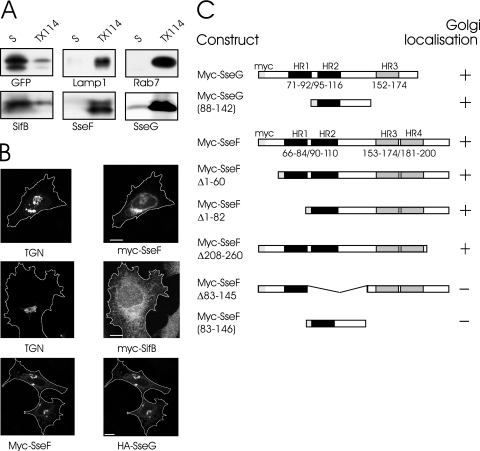
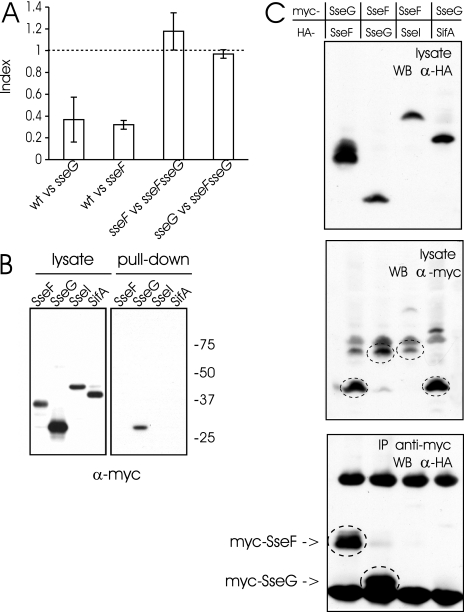

References
-
- Abrahams, G. L., and M. Hensel. 2006. Manipulating cellular transport and immune responses: dynamic interactions between intracellular Salmonella enterica and its host cells. Cell Microbiol. 8:728-737. - PubMed
-
- Beuzon, C. R., and D. W. Holden. 2001. Use of mixed infections with Salmonella strains to study virulence genes and their interactions in vivo. Microbes Infect. 3:1345-1352. - PubMed
-
- Beuzon, C. R., S. P. Salcedo, and D. W. Holden. 2002. Growth and killing of a Salmonella enterica serovar Typhimurium sifA mutant strain in the cytosol of different host cell lines. Microbiology 148:2705-2715. - PubMed
-
- Bordier, C. 1981. Phase separation of integral membrane proteins in Triton X-114 solution. J. Biol. Chem. 256:1604-1607. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
