

Slx9p facilitates efficient ITS1 processing of pre-rRNA in Saccharomyces cerevisiae
- PMID: 17018574
- PMCID: PMC1624901
- DOI: 10.1261/rna.159406
Slx9p facilitates efficient ITS1 processing of pre-rRNA in Saccharomyces cerevisiae
Abstract
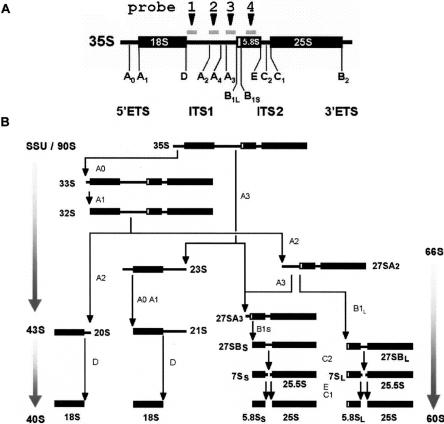
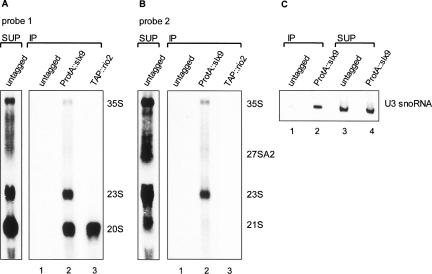
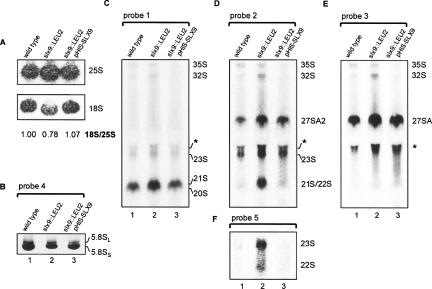
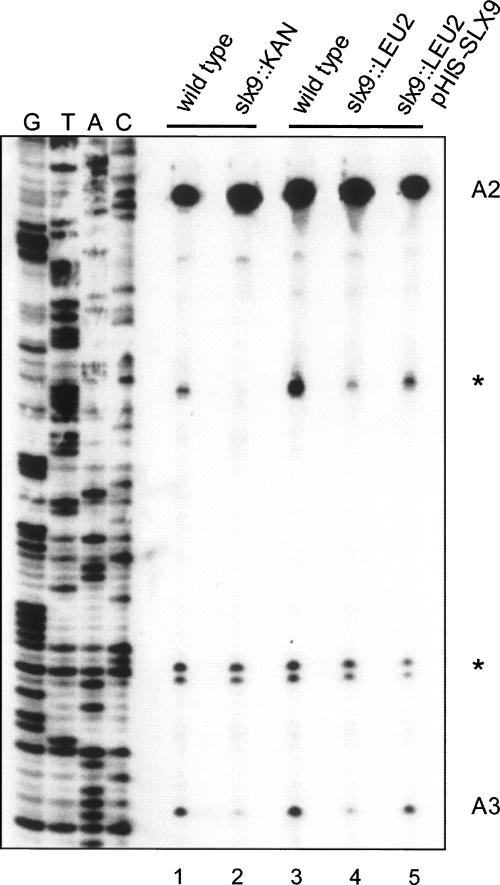
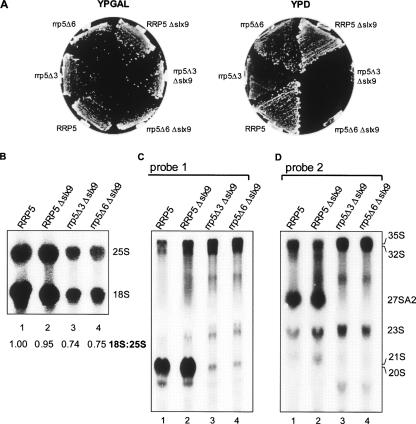
Slx9p (Ygr081cp) is a nonessential yeast protein previously linked genetically with the DNA helicase Sgs1p. Here we report that Slx9p is involved in ribosome biogenesis in the yeast Saccharomyces cerevisiae. Deletion of SLX9 results in a mild growth defect and a reduction in the level of 18S rRNA. Co-immunoprecipitation experiments showed that Slx9p is associated with 35S, 23S, and 20S pre-rRNA, as well as U3 snoRNA and, thus, is a bona fide component of pre-ribosomes. The most striking effects on pre-rRNA processing resulting from deletion of SLX9 is the accumulation of the mutually exclusive 21S and 27SA2 pre-rRNA. Furthermore, deletion of SLX9 is synthetically lethal with mutations in Rrp5p that block cleavage at either site A2 or A3. We conclude that Slx9p has a unique role in the processing events responsible for separating the 66S and 43S pre-ribosomal particles. Interestingly, homologs of Slx9p were found only in other yeast species, indicating that the protein has been considerably less well conserved during evolution than the majority of trans-acting processing factors.
Figures

Similar articles
-
Deletion of the three distal S1 motifs of Saccharomyces cerevisiae Rrp5p abolishes pre-rRNA processing at site A(2) without reducing the production of functional 40S subunits.Eukaryot Cell. 2004 Dec;3(6):1504-12. doi: 10.1128/EC.3.6.1504-1512.2004. Eukaryot Cell. 2004. PMID: 15590824 Free PMC article.
-
Domain III of Saccharomyces cerevisiae 25 S ribosomal RNA: its role in binding of ribosomal protein L25 and 60 S subunit formation.J Mol Biol. 2000 Feb 11;296(1):7-17. doi: 10.1006/jmbi.1999.3432. J Mol Biol. 2000. PMID: 10656814
-
U3 snoRNP and Rrp5p associate independently with Saccharomyces cerevisiae 35S pre-rRNA, but Rrp5p is essential for association of Rok1p.Nucleic Acids Res. 2004 Nov 2;32(19):5827-33. doi: 10.1093/nar/gkh904. Print 2004. Nucleic Acids Res. 2004. PMID: 15523097 Free PMC article.
-
Feedback regulation of ribosome assembly.Curr Genet. 2018 Apr;64(2):393-404. doi: 10.1007/s00294-017-0764-x. Epub 2017 Oct 11. Curr Genet. 2018. PMID: 29022131 Review.
-
Saccharomyces cerevisiae, a Powerful Model for Studying rRNA Modifications and Their Effects on Translation Fidelity.Int J Mol Sci. 2021 Jul 10;22(14):7419. doi: 10.3390/ijms22147419. Int J Mol Sci. 2021. PMID: 34299038 Free PMC article. Review.
Cited by
-
Las1 interacts with Grc3 polynucleotide kinase and is required for ribosome synthesis in Saccharomyces cerevisiae.Nucleic Acids Res. 2013 Jan;41(2):1135-50. doi: 10.1093/nar/gks1086. Epub 2012 Nov 21. Nucleic Acids Res. 2013. PMID: 23175604 Free PMC article.
-
FAM207A acts as a novel and potential biomarker in lung adenocarcinoma and shapes the immunesuppressive tumor microenvironment.Clin Exp Med. 2025 Apr 21;25(1):125. doi: 10.1007/s10238-025-01657-4. Clin Exp Med. 2025. PMID: 40259152 Free PMC article.
-
The Noc-domain containing C-terminus of Noc4p mediates both formation of the Noc4p-Nop14p submodule and its incorporation into the SSU processome.PLoS One. 2009 Dec 18;4(12):e8370. doi: 10.1371/journal.pone.0008370. PLoS One. 2009. PMID: 20019888 Free PMC article.
-
A screen for synthetic genetic interactions with the Saccharomyces cerevisiae hrq1ΔN allele.bioRxiv [Preprint]. 2025 Jul 4:2025.07.03.663020. doi: 10.1101/2025.07.03.663020. bioRxiv. 2025. PMID: 40631315 Free PMC article. Preprint.
-
Impact of two neighbouring ribosomal protein clusters on biogenesis factor binding and assembly of yeast late small ribosomal subunit precursors.PLoS One. 2019 Jan 17;14(1):e0203415. doi: 10.1371/journal.pone.0203415. eCollection 2019. PLoS One. 2019. PMID: 30653518 Free PMC article.
References
-
- Bennett, R.J., Sharp, J.A., Wang, J.C. Purification and characterization of the Sgs1 DNA helicase activity of Saccharomyces cerevisiae . J. Biol. Chem. 1998;273:9644–9650. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases