

Delayed satiety-like actions and altered feeding microstructure by a selective type 2 corticotropin-releasing factor agonist in rats: intra-hypothalamic urocortin 3 administration reduces food intake by prolonging the post-meal interval
- PMID: 17019404
- PMCID: PMC2748839
- DOI: 10.1038/sj.npp.1301214
Delayed satiety-like actions and altered feeding microstructure by a selective type 2 corticotropin-releasing factor agonist in rats: intra-hypothalamic urocortin 3 administration reduces food intake by prolonging the post-meal interval
Abstract
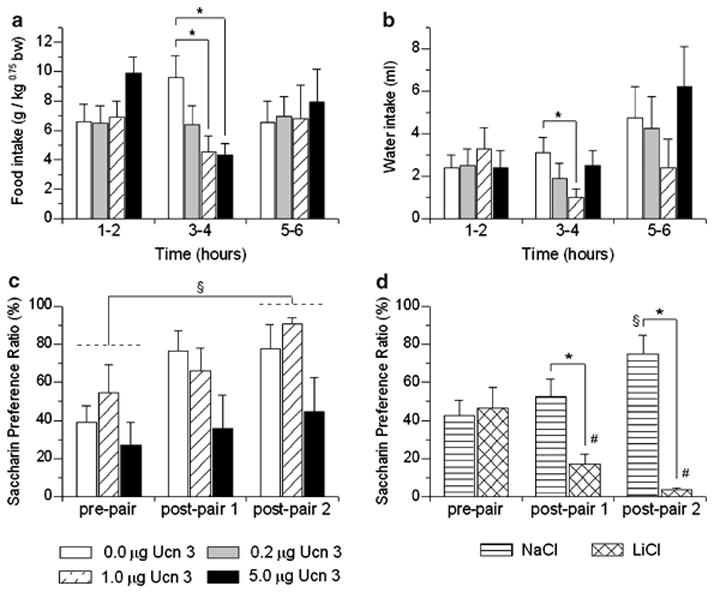
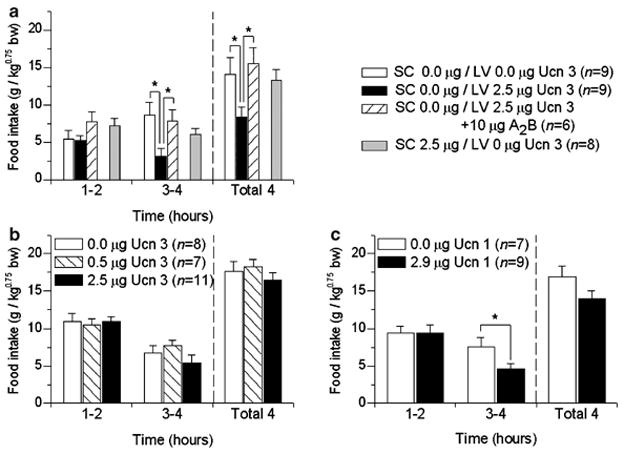
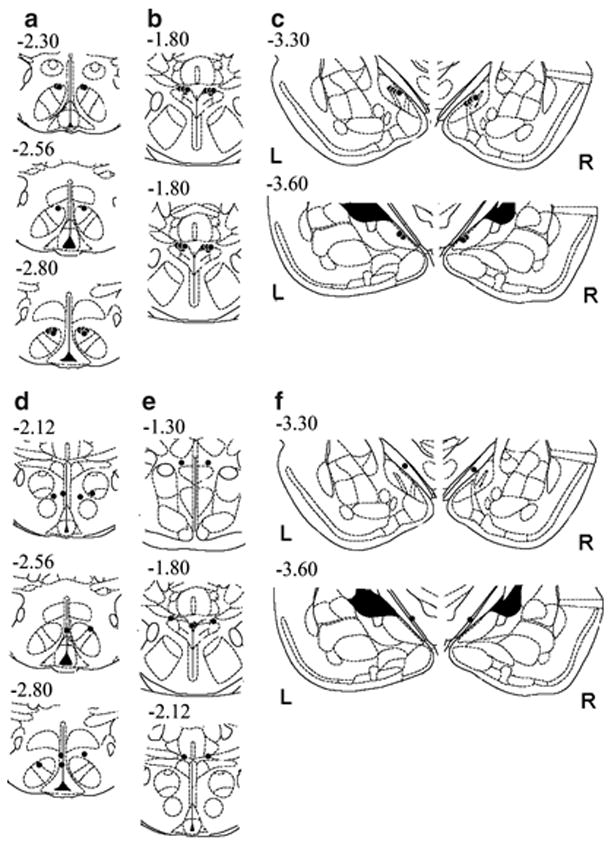
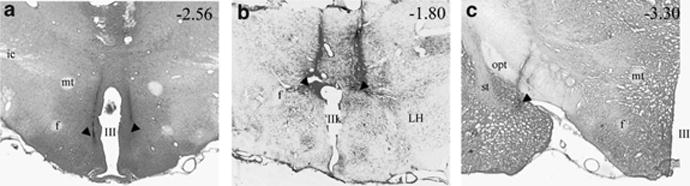
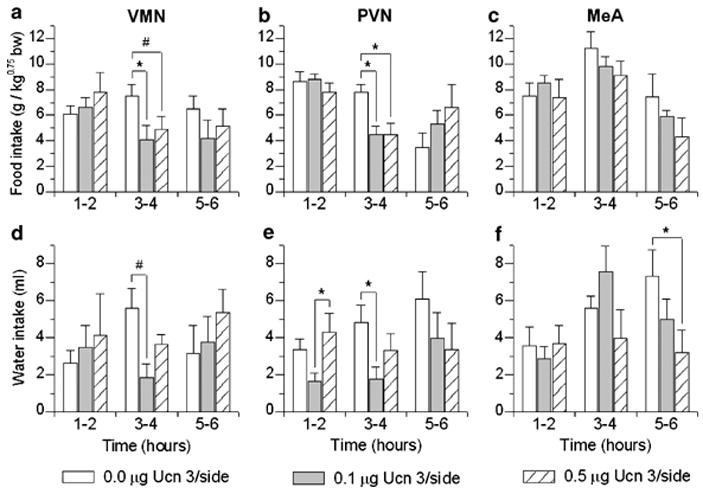
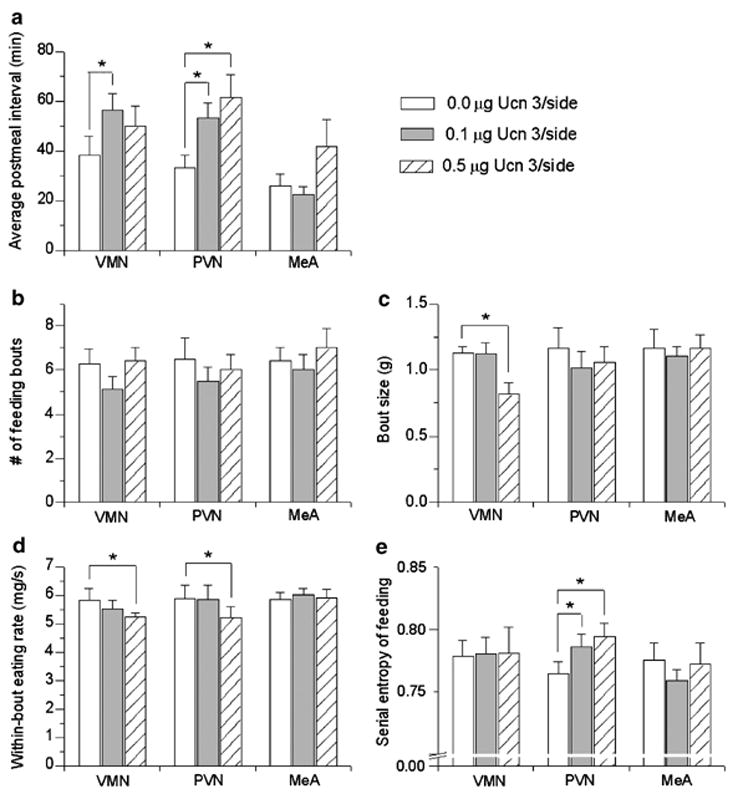
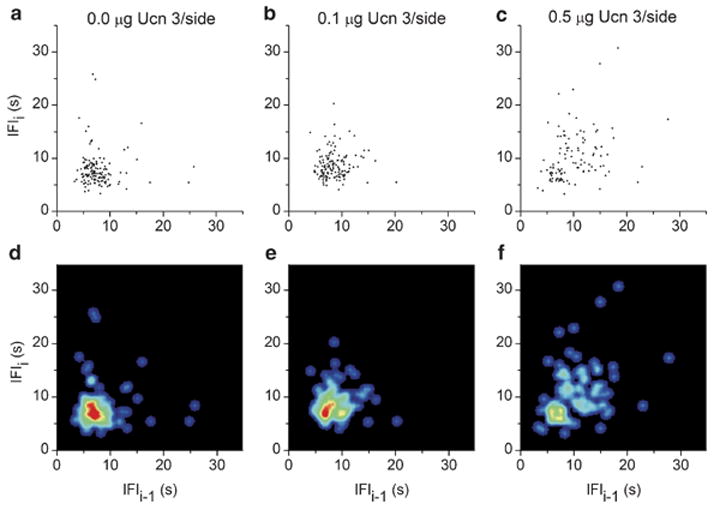
Brain corticotropin-releasing factor/urocortin (CRF/Ucn) systems are hypothesized to control feeding, with central administration of 'type 2' urocortins producing delayed anorexia. The present study sought to identify the receptor subtype, brain site, and behavioral mode of action through which Ucn 3 reduces nocturnal food intake in rats. Non-food-deprived male Wistar rats (n=176) were administered Ucn 3 into the lateral (LV) or fourth ventricle, or into the ventromedial or paraventricular nuclei of the hypothalamus (VMN, PVN) or the medial amygdala (MeA), regions in which Ucn 3 is expressed in proximity to CRF(2) receptors. LV Ucn 3 suppressed ingestion during the third-fourth post-injection hours. LV Ucn 3 anorexia was reversed by cotreatment with astressin(2)-B, a selective CRF(2) antagonist and not observed following equimole subcutaneous or fourth ventricle administration. Bilateral intra-VMN and intra-PVN infusion, more potently than LV infusion, reduced the quantity (57-73%) and duration of ingestion (32-68%) during the third-fourth post-infusion hours. LV, intra-PVN and intra-VMN infusion of Ucn 3 slowed the eating rate and reduced intake by prolonging the post-meal interval. Intra-VMN Ucn 3 reduced feeding bout size, and intra-PVN Ucn 3 reduced the regularity of eating from pellet to pellet. Ucn 3 effects were behaviorally specific, because minimal effective anorectic Ucn 3 doses did not alter drinking rate or promote a conditioned taste aversion, and site-specific, because intra-MeA Ucn 3 produced a nibbling pattern of more, but smaller meals without altering total intake. The results implicate the VMN and PVN of the hypothalamus as sites for Ucn 3-CRF(2) control of food intake.
Figures
Similar articles
-
Systemic urocortin 2, but not urocortin 1 or stressin 1-A, suppresses feeding via CRF2 receptors without malaise and stress.Br J Pharmacol. 2011 Dec;164(8):1959-75. doi: 10.1111/j.1476-5381.2011.01512.x. Br J Pharmacol. 2011. PMID: 21627635 Free PMC article.
-
Human urocortin 2, a corticotropin-releasing factor (CRF)2 agonist, and ovine CRF, a CRF1 agonist, differentially alter feeding and motor activity.J Pharmacol Exp Ther. 2004 Sep;310(3):1027-34. doi: 10.1124/jpet.104.068676. Epub 2004 Apr 28. J Pharmacol Exp Ther. 2004. PMID: 15115804
-
Urocortin in the ventromedial hypothalamic nucleus acts as an inhibitor of feeding behavior in rats.Brain Res. 2000 Apr 7;861(1):1-7. doi: 10.1016/s0006-8993(99)02378-1. Brain Res. 2000. PMID: 10751559
-
Urocortins and corticotropin releasing factor type 2 receptors in the hypothalamus and the cardiovascular system.Peptides. 2004 Oct;25(10):1711-21. doi: 10.1016/j.peptides.2004.05.024. Peptides. 2004. PMID: 15476938 Review.
-
The neurobiology of urocortin.Regul Pept. 2000 Sep 25;93(1-3):85-92. doi: 10.1016/s0167-0115(00)00180-4. Regul Pept. 2000. PMID: 11033056 Review.
Cited by
-
Small changes in meal patterns lead to significant changes in total caloric intake. Effects of diet and social status on food intake in female rhesus monkeys.Appetite. 2013 Mar;62:60-9. doi: 10.1016/j.appet.2012.11.011. Epub 2012 Nov 30. Appetite. 2013. PMID: 23207191 Free PMC article.
-
Postmeal Optogenetic Inhibition of Dorsal or Ventral Hippocampal Pyramidal Neurons Increases Future Intake.eNeuro. 2019 Jan 28;6(1):ENEURO.0457-18.2018. doi: 10.1523/ENEURO.0457-18.2018. eCollection 2019 Jan-Feb. eNeuro. 2019. PMID: 30693314 Free PMC article.
-
Salivary peptide tyrosine-tyrosine 3-36 modulates ingestive behavior without inducing taste aversion.J Neurosci. 2013 Nov 20;33(47):18368-80. doi: 10.1523/JNEUROSCI.1064-13.2013. J Neurosci. 2013. PMID: 24259562 Free PMC article.
-
Protracted withdrawal from alcohol and drugs of abuse impairs long-term potentiation of intrinsic excitability in the juxtacapsular bed nucleus of the stria terminalis.J Neurosci. 2009 Apr 29;29(17):5389-401. doi: 10.1523/JNEUROSCI.5129-08.2009. J Neurosci. 2009. PMID: 19403807 Free PMC article.
-
Urocortin 3 modulates social discrimination abilities via corticotropin-releasing hormone receptor type 2.J Neurosci. 2010 Jul 7;30(27):9103-16. doi: 10.1523/JNEUROSCI.1049-10.2010. J Neurosci. 2010. PMID: 20610744 Free PMC article.
References
-
- Behan DP, De Souza EB, Lowry PJ, Potter E, Sawchenko P, Vale WW. Corticotropin releasing factor (CRF) binding protein: a novel regulator of CRF and related peptides. Front Neuroendocrinol. 1995;16:362–382. - PubMed
-
- Borg MA, Tamborlane WV, Shulman GI, Sherwin RS. Local lactate perfusion of the ventromedial hypothalamus suppresses hypoglycemic counterregulation. Diabetes. 2003;52:663–666. - PubMed
-
- Choi S, Sparks R, Clay M, Dallman MF. Rats with hypothalamic obesity are insensitive to central leptin injections. Endocrinology. 1999;140:4426–4433. - PubMed
-
- Costa A, Poma A, Martignoni E, Nappi G, Ur E, Grossman A. Stimulation of corticotrophin-releasing hormone release by the obese (ob) gene product, leptin, from hypothalamic explants. NeuroReport. 1997;8:1131–1134. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
