

Estrogen modulation of MgATPase activity of nonmuscle myosin-II-B filaments
- PMID: 17023528
- PMCID: PMC2394735
- DOI: 10.1210/en.2006-1045
Estrogen modulation of MgATPase activity of nonmuscle myosin-II-B filaments
Abstract
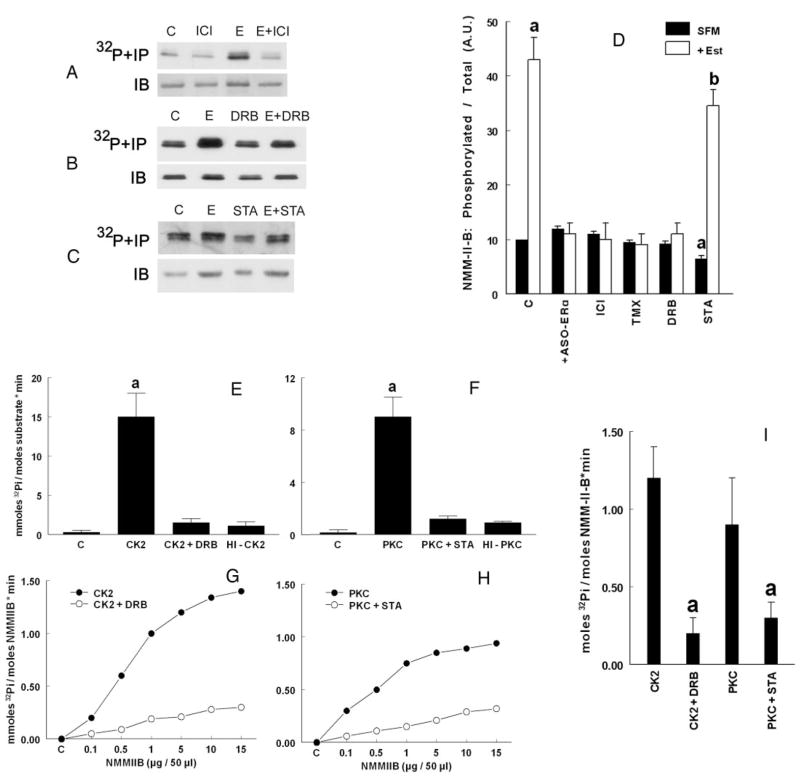
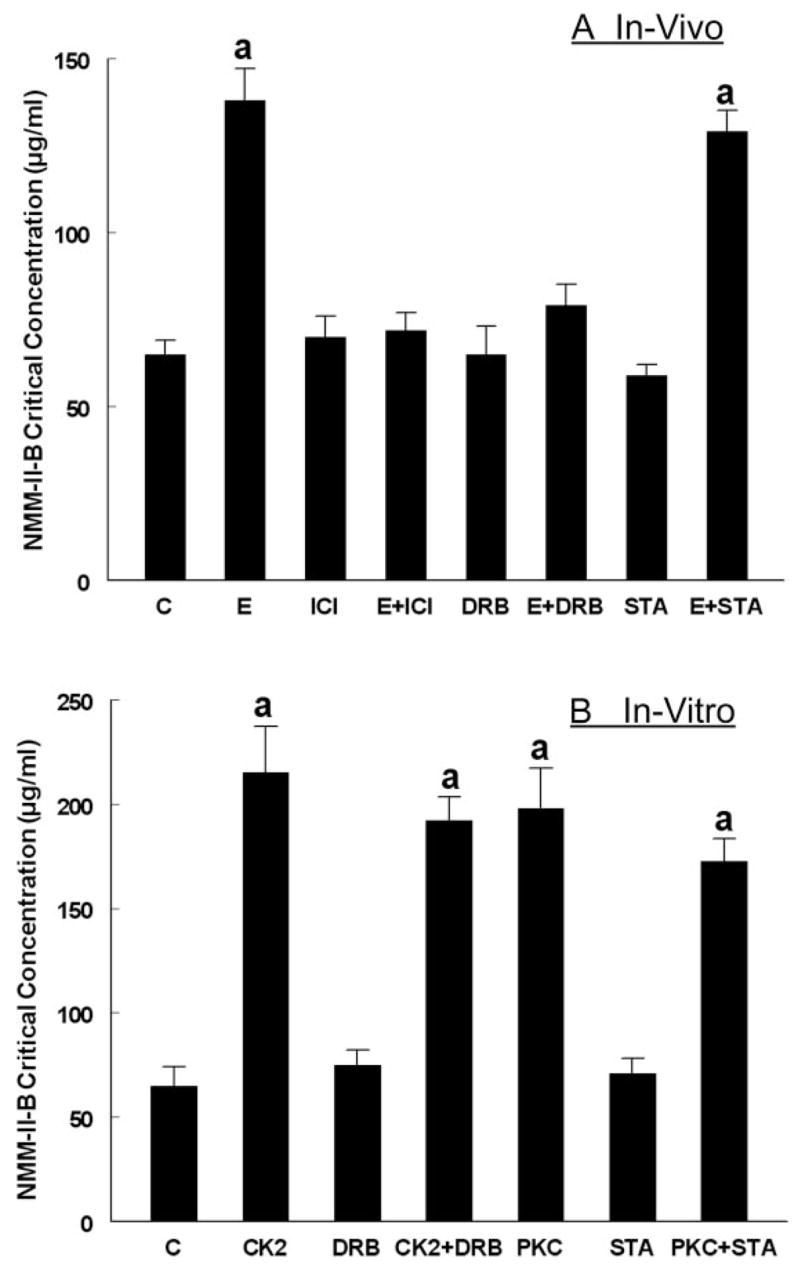
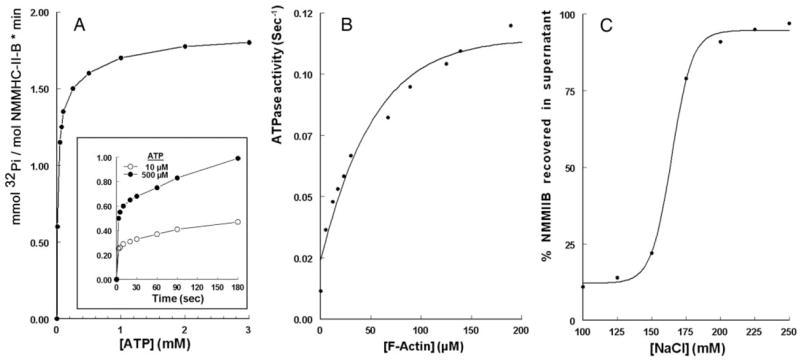
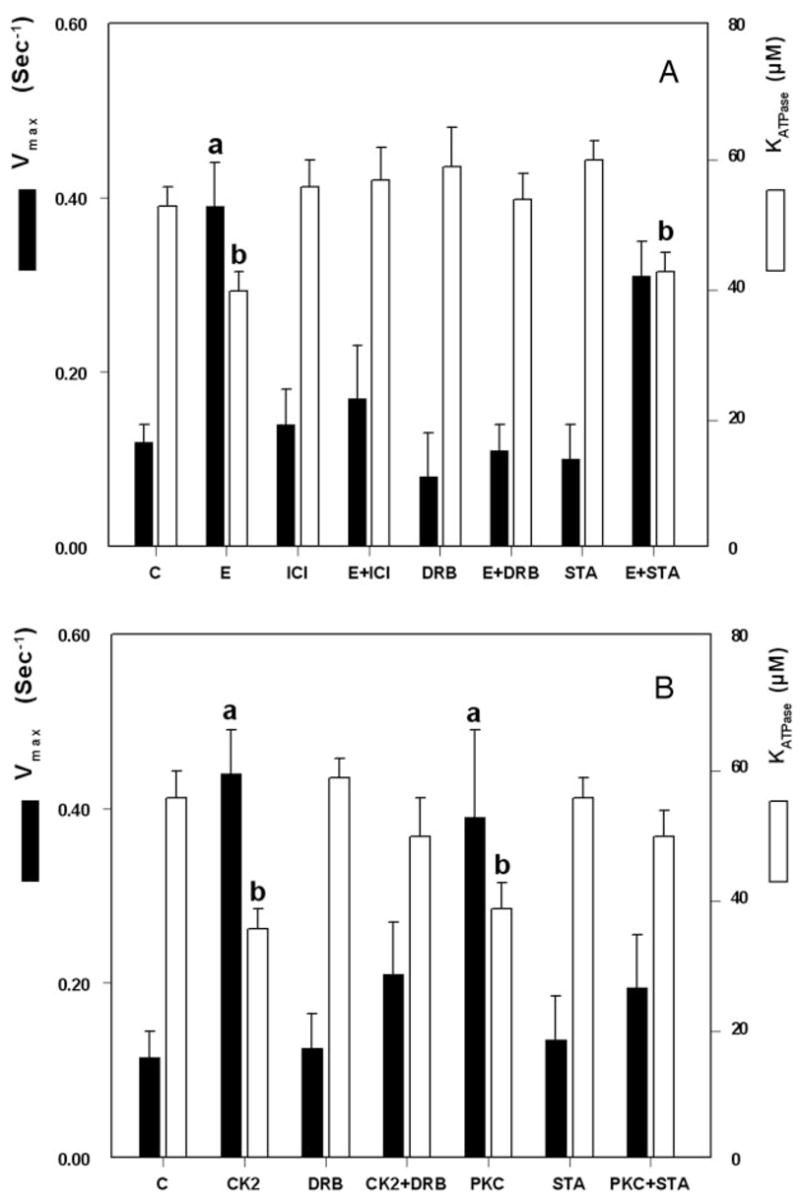
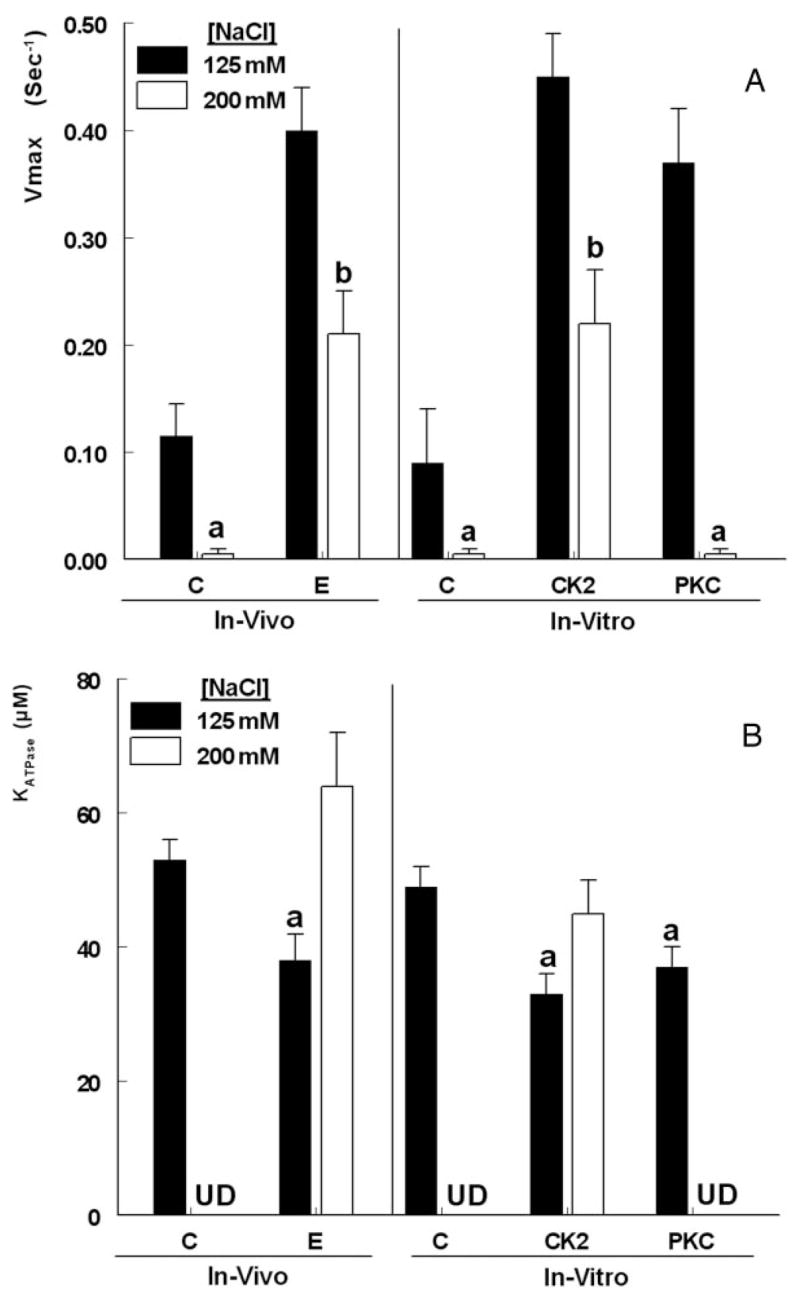
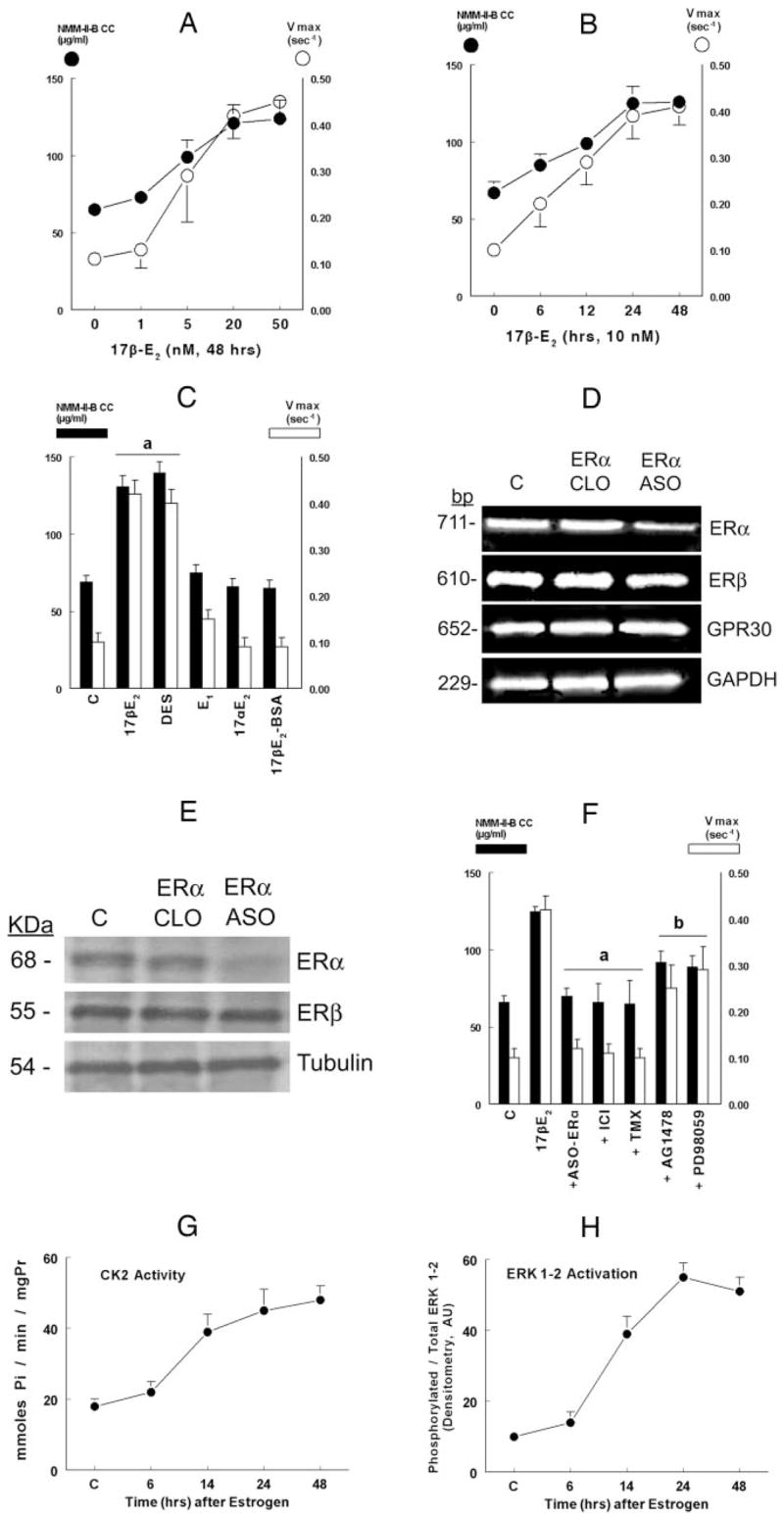
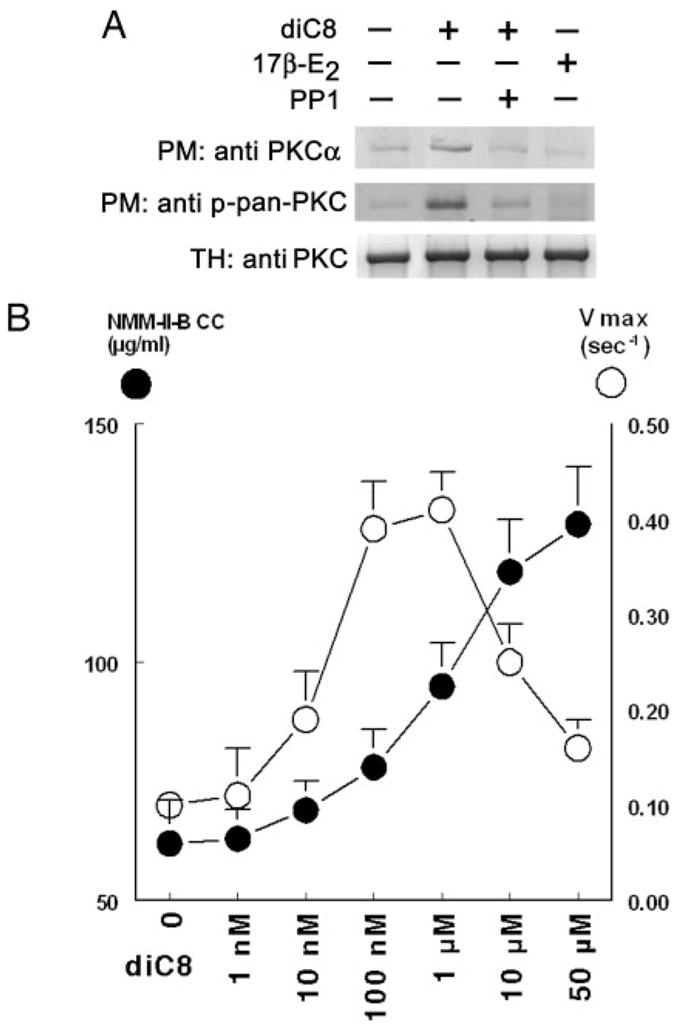
The study tested the hypothesis that estrogen controls epithelial paracellular resistance through modulation of myosin. The objective was to understand how estrogen modulates nonmuscle myosin-II-B (NMM-II-B), the main component of the cortical actomyosin in human epithelial cervical cells. Experiments used human cervical epithelial cells CaSki as a model, and end points were NMM-II-B phosphorylation, filamentation, and MgATPase activity. The results were as follows: 1) treatment with estrogen increased phosphorylation and MgATPase activity and decreased NMM-II-B filamentation; 2) estrogen effects could be blocked by antisense nucleotides for the estrogen receptor-alpha and by ICI-182,780, tamoxifen, and the casein kinase-II (CK2) inhibitor, 5,6-dichloro-1-beta-(D)-ribofuranosylbenzimidazole and attenuated by AG1478 and PD98059 (inhibitors of epithelial growth factor receptor and ERK/MAPK) but not staurosporine [blocker of protein kinase C (PKC)]; 3) treatments with the PKC activator sn-1,2-dioctanoyl diglyceride induced biphasic effect on NMM-II-B MgATPase activity: an increase at 1 nm to 1 microM and a decrease in activity at more than 1 microM; 4) sn-1,2-dioctanoyl diglyceride also decreased NMM-II-B filamentation in a monophasic and saturable dose dependence (EC(50) 1-10 microM); 5) when coincubated directly with purified NMM-II-B filaments, both CK2 and PKC decreased filamentation and increased MgATPase activity; 6) assays done on disassembled NMM-II-B filaments showed MgATPase activity in filaments obtained from estrogen-treated cells but not estrogen-depleted cells; and 7) incubations in vitro with CK2, but not PKC, facilitated MgATPase activity, even in disassembled NMM-II-B filaments. The results suggest that estrogen, in an effect mediated by estrogen receptor-alpha and CK2 and involving the epithelial growth factor receptor and ERK/MAPK cascades, increases NMM-II-B MgATPase activity independent of NMM-II-B filamentation status.
Figures
References
-
- Ussing HH, Zerahn K. Active transport of sodium as the source of electric current in the short-circuited isolated frog skin. Acta Physiol Scand. 1951;23:110–127. - PubMed
-
- Gorodeski GI. The cultured human cervical epithelium: a new model for studying transepithelial paracellular transport. J Soc Gynecol Invest. 1996;3:267–280. - PubMed
-
- Zeng R, Li X, Gorodeski GI. Estrogen abrogates transcervical tight junctional resistance by acceleration of occludin modulation. J Clin Endocrinol Metab. 2004;89:5145–5155. - PubMed
-
- Gorodeski GI. Estrogen increases the permeability of the cultured human cervical epithelium by modulating cell deformability. Am J Physiol. 1998;275:C888–C899. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
