

Effects of the trinucleotide preceding the self-cleavage site on eggplant latent viroid hammerheads: differences in co- and post-transcriptional self-cleavage may explain the lack of trinucleotide AUC in most natural hammerheads
- PMID: 17028097
- PMCID: PMC1636495
- DOI: 10.1093/nar/gkl717
Effects of the trinucleotide preceding the self-cleavage site on eggplant latent viroid hammerheads: differences in co- and post-transcriptional self-cleavage may explain the lack of trinucleotide AUC in most natural hammerheads
Abstract
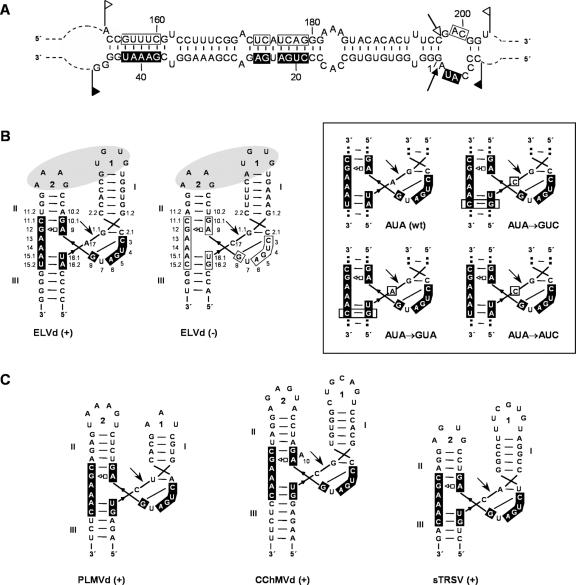
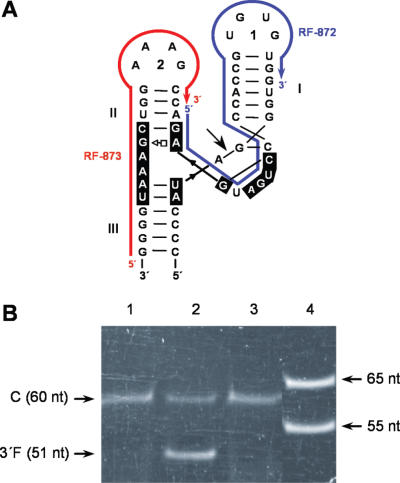
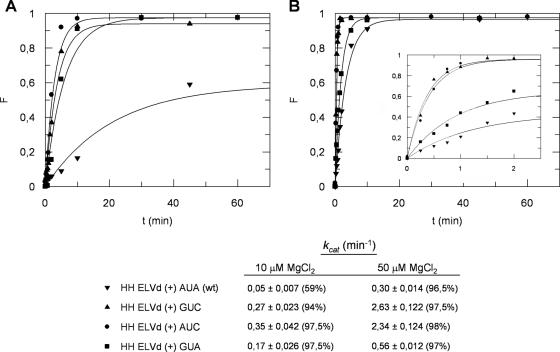
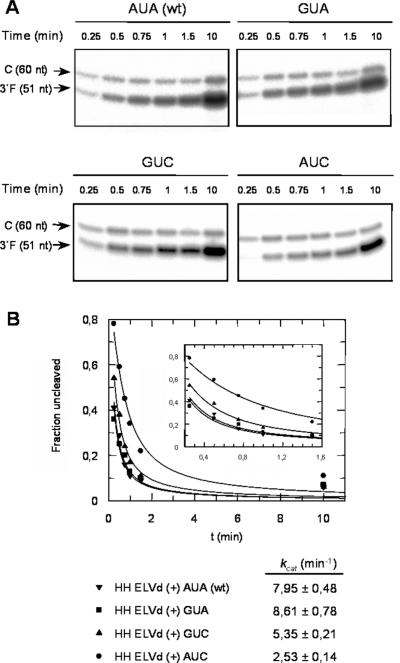
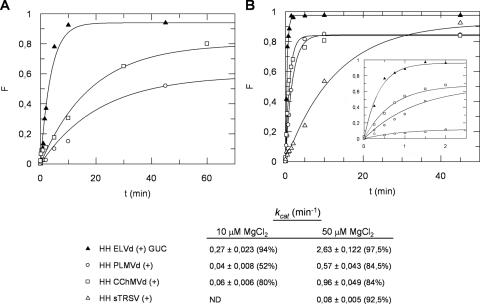
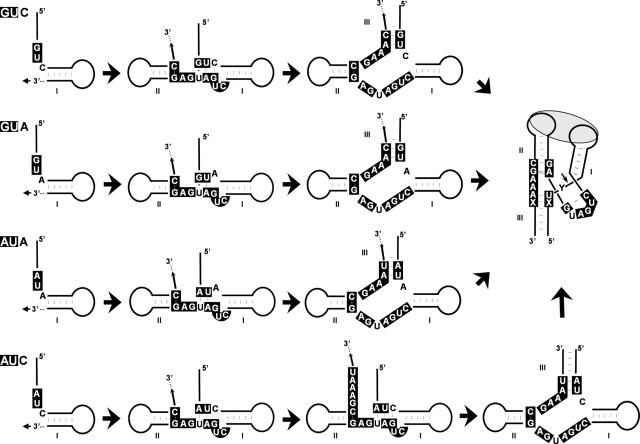
Eggplant latent viroid (ELVd) can form stable hammerhead structures in its (+) and (-) strands. These ribozymes have the longest helices I reported in natural hammerheads, with that of the ELVd (+) hammerhead being particularly stable (5/7 bp are G-C). Moreover, the trinucleotide preceding the self-cleavage site of this hammerhead is AUA, which together with GUA also found in some natural hammerheads, deviate from the GUC present in most natural hammerheads including the ELVd (-) hammerhead. When the AUA trinucleotide preceding the self-cleavage site of the ELVd (+) hammerhead was substituted by GUA and GUC, as well as by AUC (essentially absent in natural hammerheads), the values of the self-cleavage rate constants at low magnesium of the purified hammerheads were: ELVd-(+)-AUC approximately ELVd-(+)-GUC>ELVd-(+)-GUA> ELVd-(+)-AUA. However, the ELVd-(+)-AUC hammerhead was the catalytically less efficient during in vitro transcription, most likely because of the transient adoption of catalytically-inactive metastable structures. These results suggest that natural hammerheads have been evolutionary selected to function co-transcriptionally, and provide a model explaining the lack of trinucleotide AUC preceding the self-cleavage site of most natural hammerheads. Comparisons with other natural hammerheads showed that the ELVd-(+)-GUC and ELVd-(+)-AUC hammerheads are the catalytically most active in a post-transcriptional context with low magnesium.
Figures
References
-
- Prody G.A., Bakos J.T., Buzayan J.M., Schneider I.R., Bruening G. Autolytic processing of dimeric plant virus satellite RNA. Science. 1986;231:1577–1580. - PubMed
-
- Flores R., Hernández C., De la Peña M., Vera A., Daròs J.A. Hammerhead ribozyme structure and function in plant RNA replication. Methods Enzymol. 2001;341:540–552. - PubMed
-
- Flores R., Hernández C., Martínez de Alba E., Daròs J.A., Di Serio F. Viroids and viroid-host interactions. Ann. Rev. Phytopathol. 2005;43:117–139. - PubMed
-
- Forster A.C., Symons R.H. Self-cleavage of plus and minus RNAs of a virusoid and a structural model for the active sites. Cell. 1987;49:211–220. - PubMed
