

Molecular interactions between the specialist herbivore Manduca sexta (Lepidoptera, Sphingidae) and its natural host Nicotiana attenuata. VII. Changes in the plant's proteome
- PMID: 17028148
- PMCID: PMC1676057
- DOI: 10.1104/pp.106.088781
Molecular interactions between the specialist herbivore Manduca sexta (Lepidoptera, Sphingidae) and its natural host Nicotiana attenuata. VII. Changes in the plant's proteome
Abstract
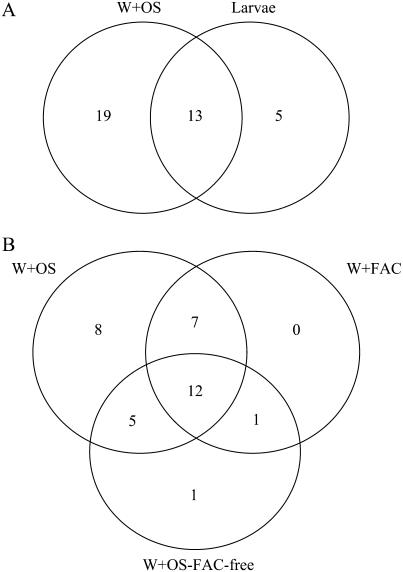
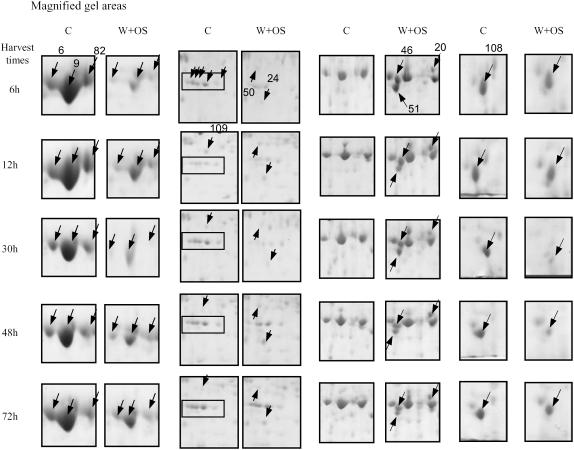
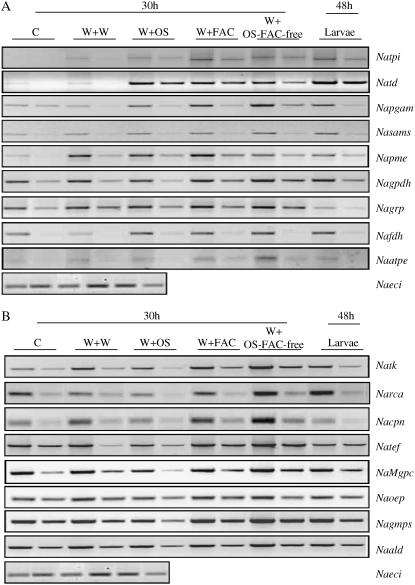
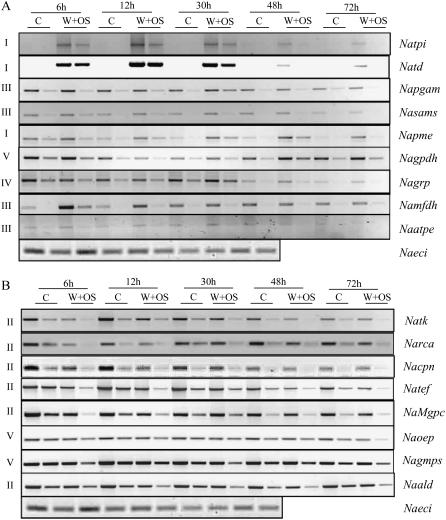
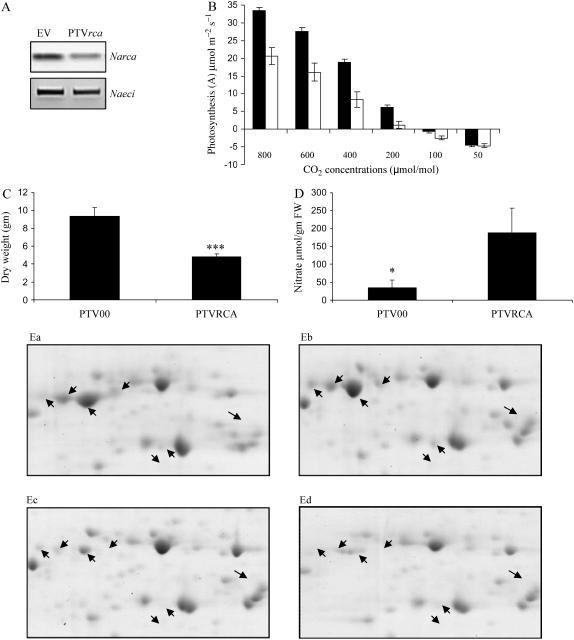
When Manduca sexta attacks Nicotiana attenuata, fatty acid-amino acid conjugates (FACs) in the larvae's oral secretions (OS) are introduced into feeding wounds. These FACs trigger a transcriptional response that is similar to the response induced by insect damage. Using two-dimensional gel electrophoresis, matrix-assisted laser desorption ionization-time of flight, and liquid chromatography-tandem mass spectrometry, we characterized the proteins in phenolic extracts and in a nuclear fraction of leaves elicited by larval attack, and/or in leaves wounded and treated with OS, FAC-free OS, and synthetic FACs. Phenolic extracts yielded approximately 600 protein spots, many of which were altered by elicitation, whereas nuclear protein fractions yielded approximately 100 spots, most of which were unchanged by elicitation. Reproducible elicitor-induced changes in 90 spots were characterized. In general, proteins that increased were involved in primary metabolism, defense, and transcriptional and translational regulation; those that decreased were involved in photosynthesis. Like the transcriptional defense responses, proteomic changes were strongly elicited by the FACs in OS. A semiquantitative reverse transcription-PCR approach based on peptide sequences was used to compare transcript and protein accumulation patterns for 17 candidate proteins. In six cases the patterns of elicited transcript accumulation were consistent with those of elicited protein accumulation. Functional analysis of one of the identified proteins involved in photosynthesis, RuBPCase activase, was accomplished by virus-induced gene silencing. Plants with decreased levels of RuBPCase activase protein had reduced photosynthetic rates and RuBPCase activity, and less biomass, responses consistent with those of herbivore-attacked plants. We conclude that the response of the plant's proteome to herbivore elicitation is complex, and integrated transcriptome-proteome-metabolome analysis is required to fully understand this ubiquitous ecological interaction.
Figures



References
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
