

The tomato NBARC-LRR protein Prf interacts with Pto kinase in vivo to regulate specific plant immunity
- PMID: 17028203
- PMCID: PMC1626632
- DOI: 10.1105/tpc.106.044016
The tomato NBARC-LRR protein Prf interacts with Pto kinase in vivo to regulate specific plant immunity
Abstract
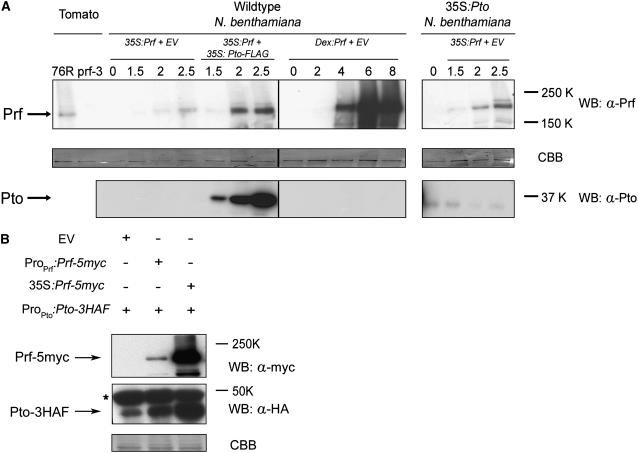
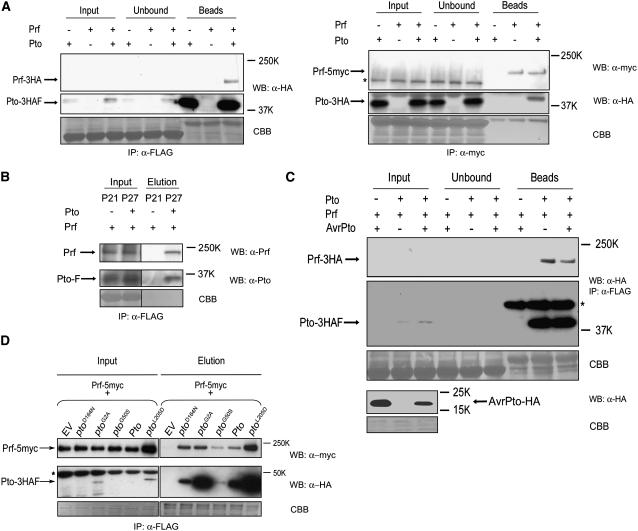
Immunity in tomato (Solanum lycopersicum) to Pseudomonas syringae bacteria expressing the effector proteins AvrPto and AvrPtoB requires both Pto kinase and the NBARC-LRR (for nucleotide binding domain shared by Apaf-1, certain R gene products, and CED-4 fused to C-terminal leucine-rich repeats) protein Prf. Pto plays a direct role in effector recognition within the host cytoplasm, but the role of Prf is unknown. We show that Pto and Prf are coincident in the signal transduction pathway that controls ligand-independent signaling. Pto and Prf associate in a coregulatory interaction that requires Pto kinase activity and N-myristoylation for signaling. Pto interacts with a unique Prf N-terminal domain outside of the NBARC-LRR domain and resides in a high molecular weight recognition complex dependent on the presence of Prf. In this complex, both Pto and Prf contribute to specific recognition of AvrPtoB. The data suggest that the role of Pto is confined to the regulation of Prf and that the bacterial effectors have evolved to target this coregulatory molecular switch.
Figures





References
-
- Aoyama, T., and Chua, N.-H. (1997). A glucocorticoid-mediated transcriptional induction system in transgenic plants. Plant J. 11 605–612. - PubMed
-
- Axtell, M.J., and Staskawicz, B. (2003). Initiation of RPS2-specified disease resistance in Arabidopsis is coupled to the AvrRpt2-directed elimination of RIN4. Cell 112 369–377. - PubMed
-
- Ballvora, A., Ercolano, M., Weiss, J., Meksem, K., Bormann, C., Oberhagemann, P., Salamini, F., and Gebhardt, C. (2002). The R1 gene for potato resistance to late blight (Phytophthora infestans) belongs to the leucine zipper/NBS/LRR class of plant resistance genes. Plant J. 30 361–371. - PubMed
-
- Belkhadir, Y., Subramaniam, R., and Dangl, J.L. (2004). Plant disease resistance protein signaling: NBS-LRR proteins and their partners. Curr. Opin. Plant Biol. 7 391–399. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
