

Drought, abscisic acid and transpiration rate effects on the regulation of PIP aquaporin gene expression and abundance in Phaseolus vulgaris plants
- PMID: 17028296
- PMCID: PMC2803586
- DOI: 10.1093/aob/mcl219
Drought, abscisic acid and transpiration rate effects on the regulation of PIP aquaporin gene expression and abundance in Phaseolus vulgaris plants
Abstract
Background and aims: Drought causes a decline of root hydraulic conductance, which aside from embolisms, is governed ultimately by aquaporins. Multiple factors probably regulate aquaporin expression, abundance and activity in leaf and root tissues during drought; among these are the leaf transpiration rate, leaf water status, abscisic acid (ABA) and soil water content. Here a study is made of how these factors could influence the response of aquaporin to drought.
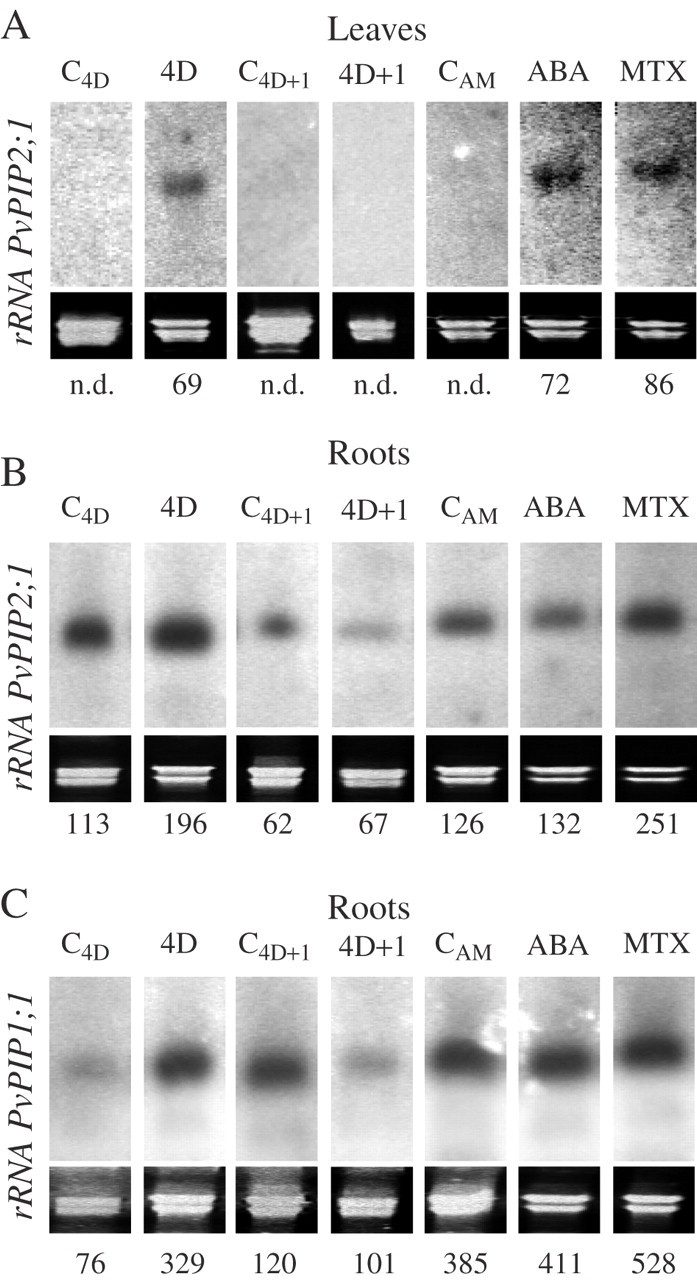
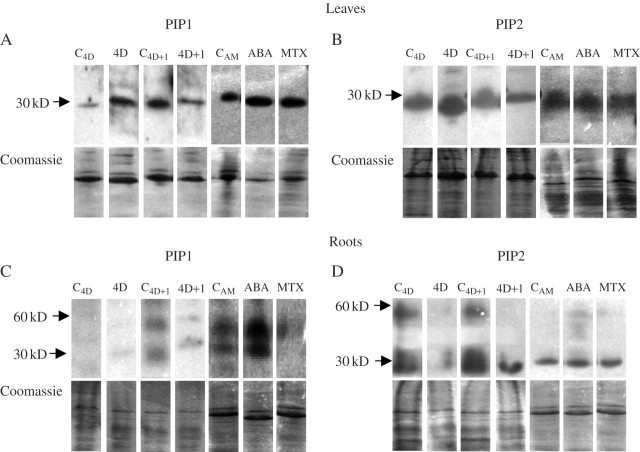
Methods: Three plasma membrane intrinsic proteins (PIPs) or aquaporins were cloned from Phaseolus vulgaris plants and their expression was analysed after 4 d of water deprivation and also 1 d after re-watering. The effects of ABA and of methotrexate (MTX), an inhibitor of stomatal opening, on gene expression and protein abundance were also analysed. Protein abundance was examined using antibodies against PIP1 and PIP2 aquaporins. At the same time, root hydraulic conductance (L), transpiration rate, leaf water status and ABA tissue concentration were measured.
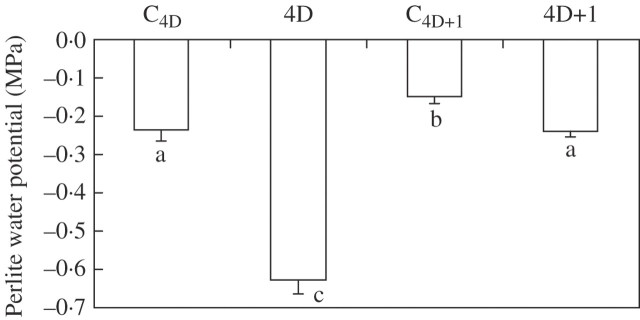
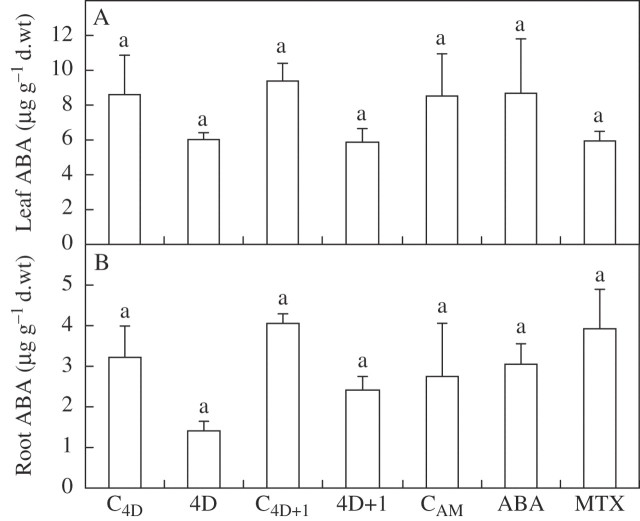
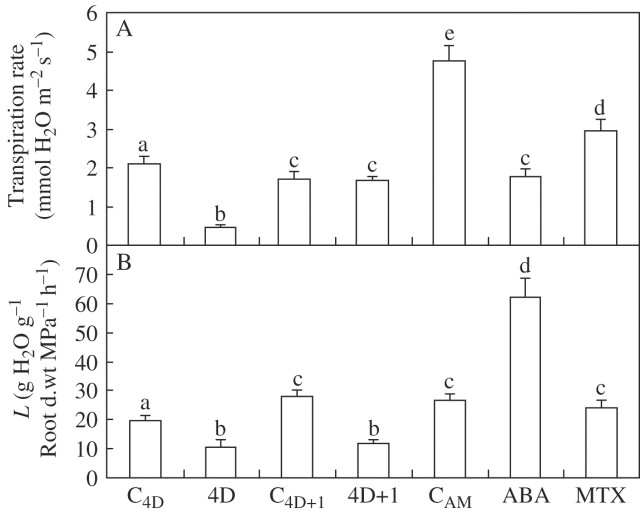
Key results: None of the treatments (drought, ABA or MTX) changed the leaf water status or tissue ABA concentration. The three treatments caused a decline in the transpiration rate and raised PVPIP2;1 gene expression and PIP1 protein abundance in the leaves. In the roots, only the drought treatment raised the expression of the three PIP genes examined, while at the same time diminishing PIP2 protein abundance and L. On the other hand, ABA raised both root PIP1 protein abundance and L.
Conclusions: The rise of PvPIP2;1 gene expression and PIP1 protein abundance in the leaves of P. vulgaris plants subjected to drought was correlated with a decline in the transpiration rate. At the same time, the increase in the expression of the three PIP genes examined caused by drought and the decline of PIP2 protein abundance in the root tissues were not correlated with any of the parameters measured.
Figures

References
-
- Alexandersson E, Fraysse L, Sjövall-Larsen S, Gustavsson S, Fellert M, Karlsson M, et al. (2005) Whole gene family expression and drought stress regulation of aquaporins. Plant Molecular Biology 59469–484. - PubMed
-
- Aroca R. (2006) Exogenous catalase and ascorbate modify the effects of abscisic acid (ABA) on root hydraulic properties in Phaseolus vulgaris L. plants. Journal of Plant Growth Regulation 2510–17.
-
- Aroca R, Vernieri P, Irigoyen JJ, Sánchez-Díaz M, Tognoni F, Pardossi A. (2003) Involvement of abscisic acid in leaf and root of maize (Zea mays L.) in avoiding chilling-induced water stress. Plant Science 165671–679.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
