

CD36 is important for chylomicron formation and secretion and may mediate cholesterol uptake in the proximal intestine
- PMID: 17030189
- PMCID: PMC1994908
- DOI: 10.1053/j.gastro.2006.08.012
CD36 is important for chylomicron formation and secretion and may mediate cholesterol uptake in the proximal intestine
Abstract
Background & aims: Studies are aimed to determine the role of CD36 in intestinal lipid absorption.
Methods: Knock-out (KO) and wild-type (WT) lymph fistula mice were used to study fatty acids (FA) and cholesterol uptake, and chylomicron formation and secretion. Uptake of FA and cholesterol was studied by using sucrose polybehenate and fecal dual isotope methods, respectively.
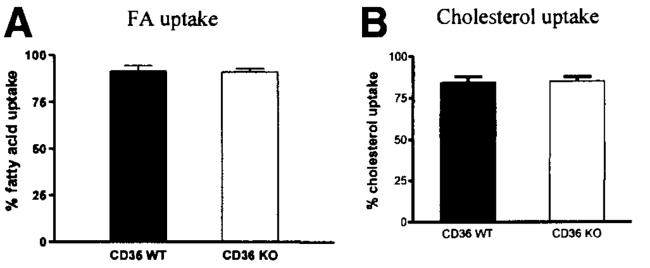
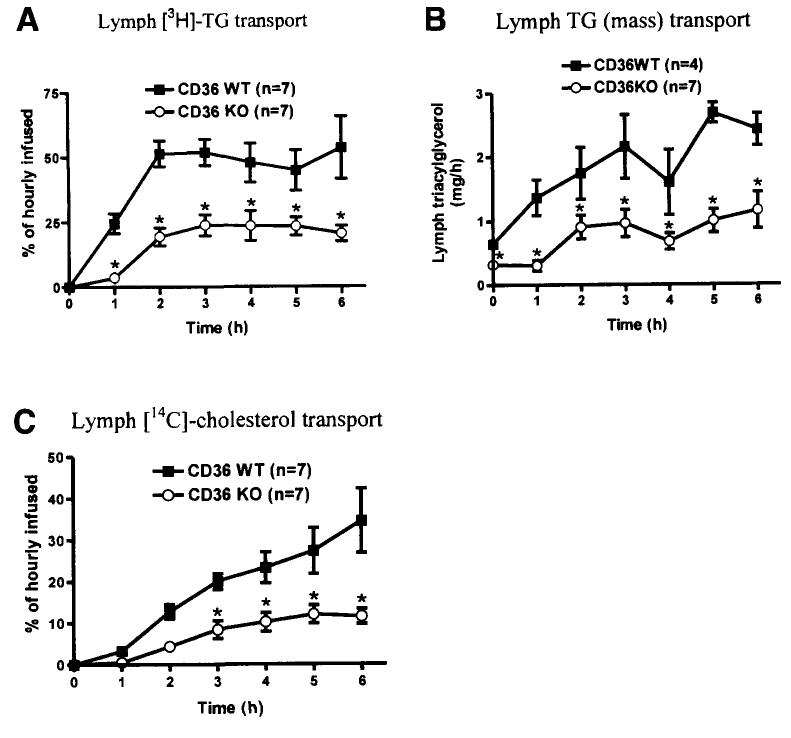
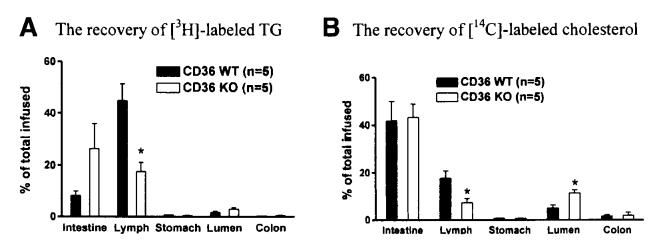
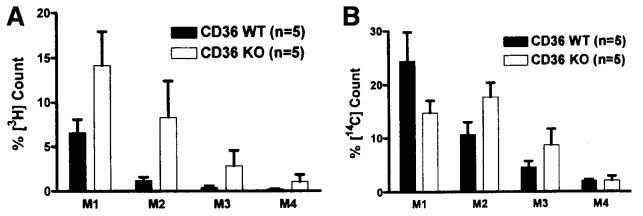
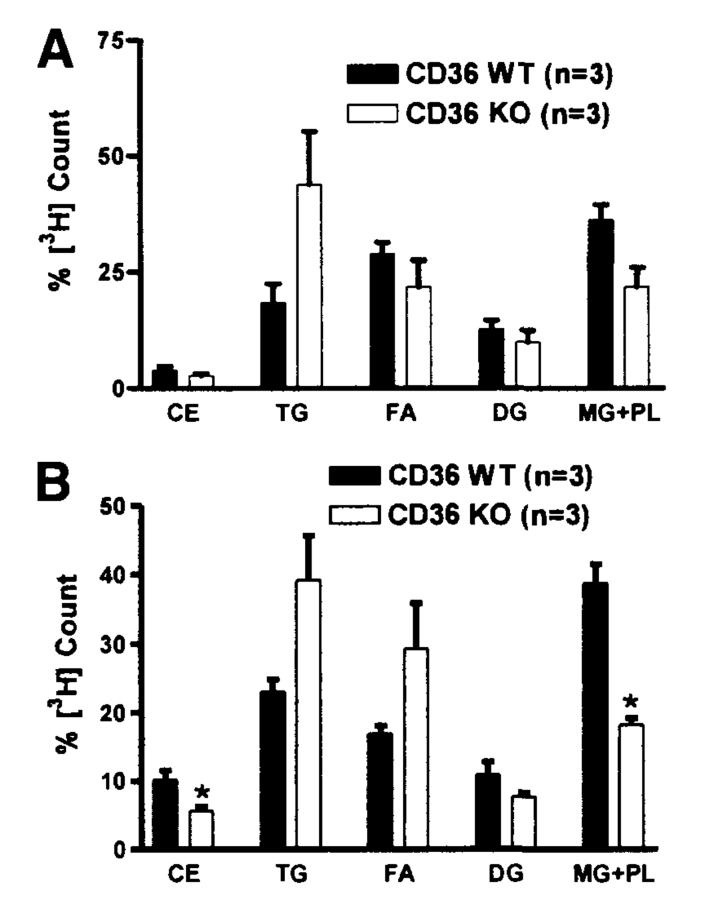
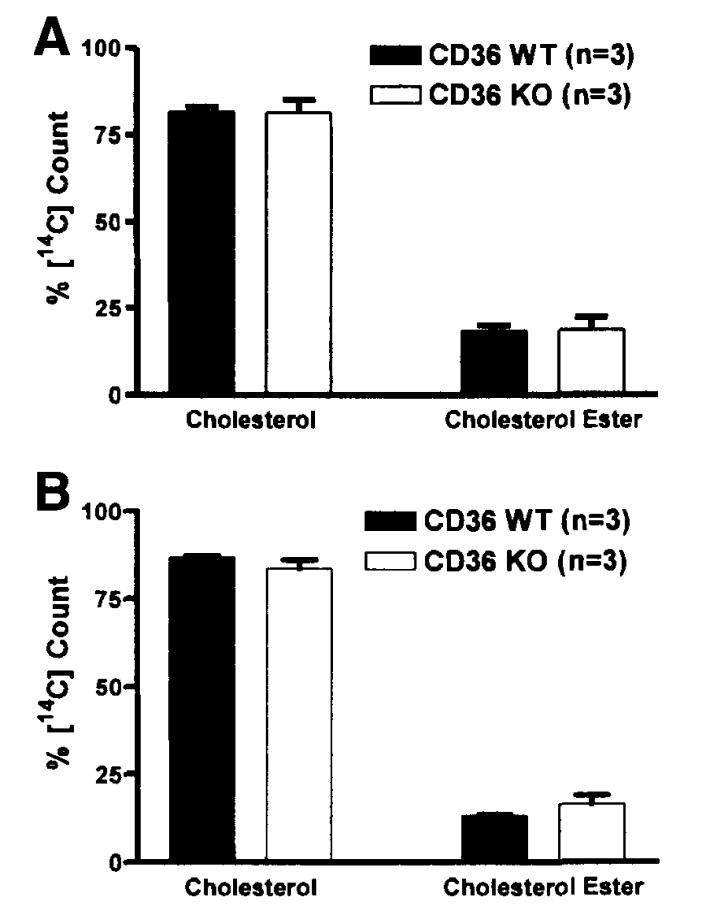
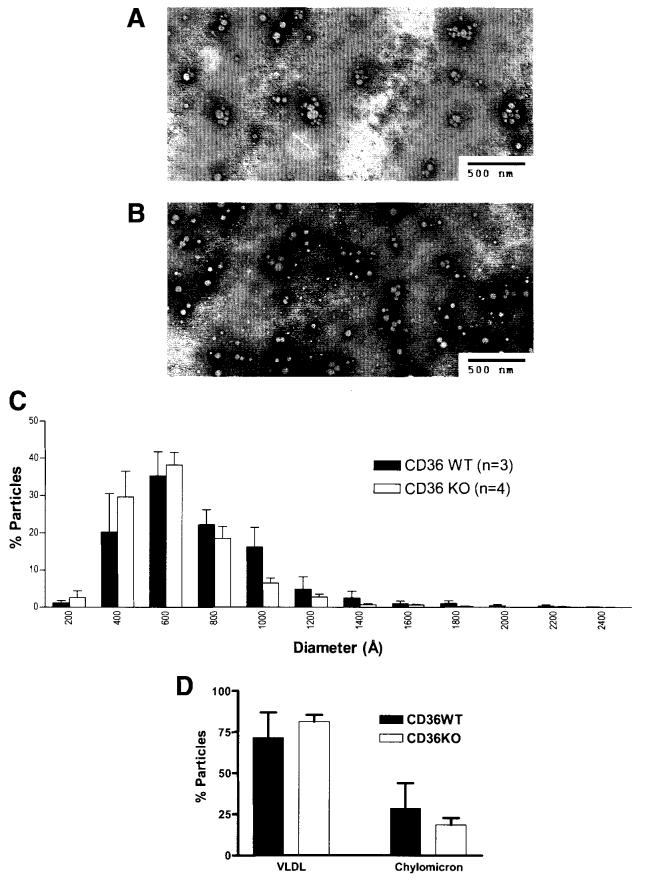
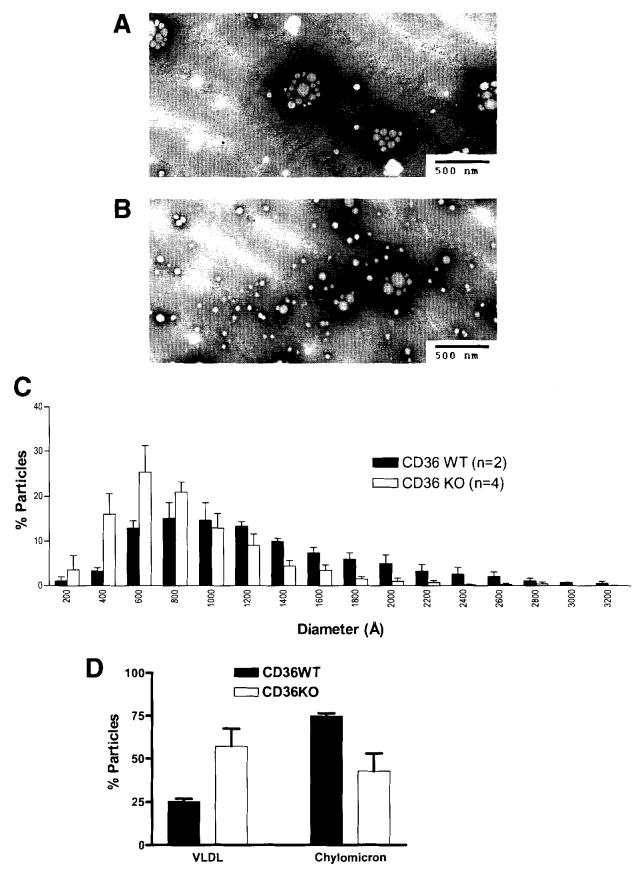
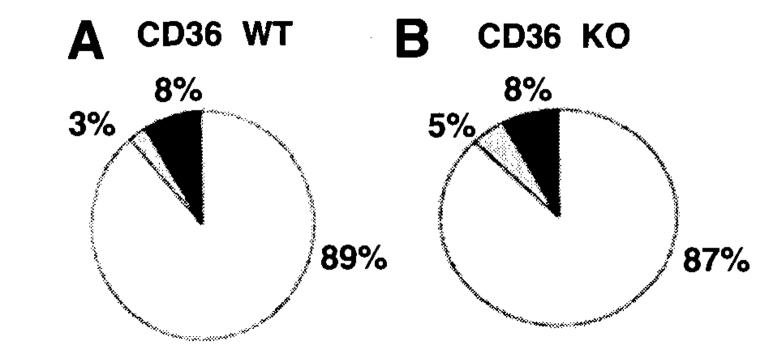
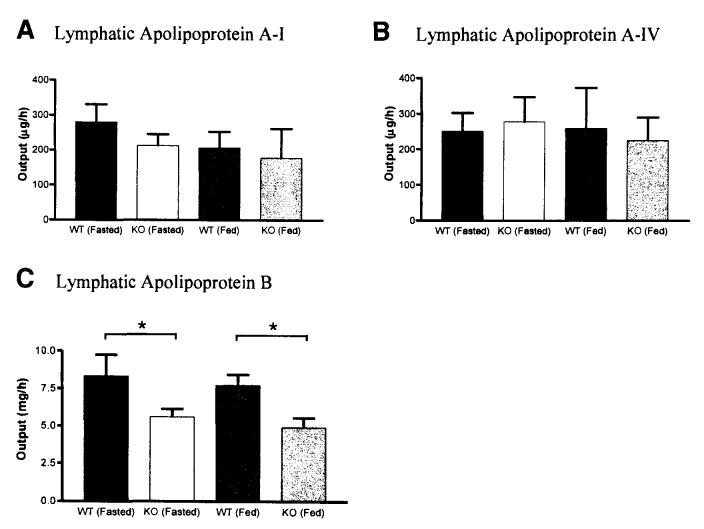
Results: The CD36 KO exhibited significant accumulation of dietary cholesterol in the intestinal lumen at the end of 6-hour lipid infusion and significant reduction of dietary cholesterol transport into the lymph. Fecal dual isotope studies, however, did not show any significant difference in cholesterol uptake, suggesting that given sufficient time, the KO intestine could compensate for the reduced cholesterol uptake observed in the acute lymph fistula studies. Recovery of dietary FA in the intestinal lumen was comparable between WT and KO, consistent with the sucrose polybehenate study. However, the KO mice accumulated more, albeit not significantly, dietary triacylglycerols in the intestine, followed by a significant reduction in lymphatic transport. The ratio of intestinal dietary triacylglycerols to FA was not higher in WT than KO, arguing against impaired lipid esterification. It is rather a deficiency in the formation and secretion of chylomicrons, as supported by the significantly less apolipoprotein B-48 and the smaller, albeit not significantly, lipoprotein particles secreted into the lymph of the KO.
Conclusions: CD36 may play an important role in chylomicron formation and secretion and may also facilitate cholesterol uptake in the proximal intestine.
Figures
References
-
- Phillips DR, Agin PP. Platelet plasma membrane glycoproteins. Evidence for the presence of nonequivalent disulfide bonds using nonreduced-reduced two-dimensional gel electrophoresis. J Biol Chem. 1977;252:2121–2126. - PubMed
-
- Talle MA, Rao PE, Westberg E, Allegar N, Makowski M, Mittler RS, Goldstein G. Patterns of antigenic expression on human monocytes as defined by monoclonal antibodies. Cell Immunol. 1983;78:83–99. - PubMed
-
- Knowles DM, Tolidjian B, Marboe C, D'Agati V, Grimes M, Chess L. Monoclonal anti-human monocyte antibodies OKM1 and OKM5 possess distinctive tissue distributions including differential reactivity with vascular endothelium. J Immunol. 1984;132:2170–2173. - PubMed
-
- Edelman P, Vinci G, Villeval JL, Vainchenker W, Henri A, Miglierina R, Rouger P, Reviron J, Breton-Gorius J, Sureau C. A monoclonal antibody against an erythrocyte ontogenic antigen identifies fetal and adult erythroid progenitors. Blood. 1986;67:56–63. - PubMed
-
- Abumrad NA, el Maghrabi MR, Amri EZ, Lopez E, Grimaldi PA. Cloning of a rat adipocyte membrane protein implicated in binding or transport of long-chain fatty acids that is induced during preadipocyte differentiation. Homology with human CD36. J Biol Chem. 1993;268:17665–17668. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
