

High-molecular-mass APOBEC3G complexes restrict Alu retrotransposition
- PMID: 17030807
- PMCID: PMC1592537
- DOI: 10.1073/pnas.0604524103
High-molecular-mass APOBEC3G complexes restrict Alu retrotransposition
Abstract
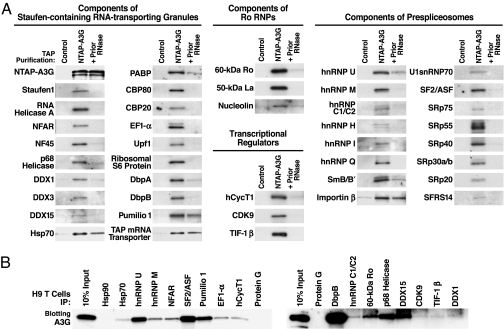
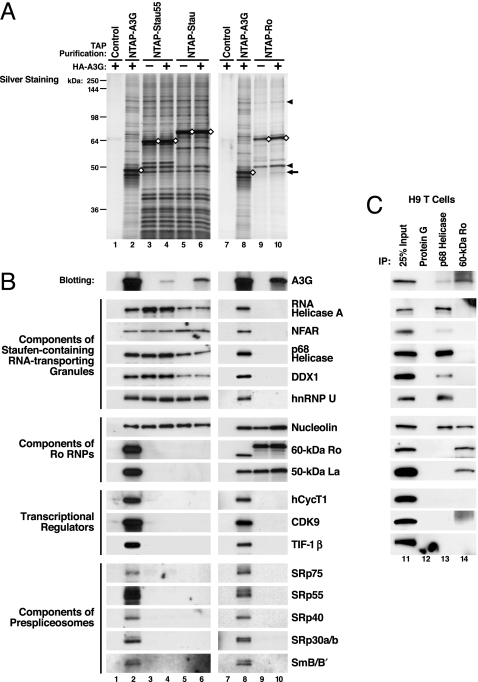
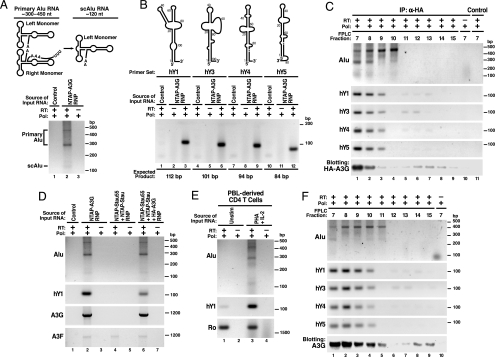
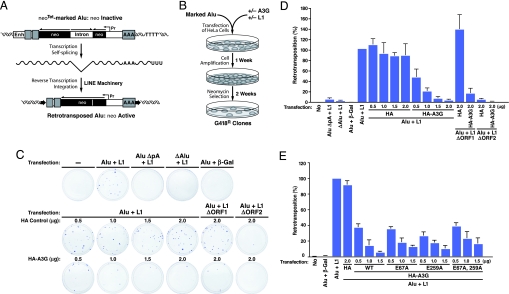
APOBEC3G (A3G) and related deoxycytidine deaminases are potent intrinsic antiretroviral factors. A3G is expressed either as an enzymatically active low-molecular-mass (LMM) form or as an enzymatically inactive high-molecular-mass (HMM) ribonucleoprotein complex. Resting CD4 T cells exclusively express LMM A3G, where it functions as a powerful postentry restriction factor for HIV-1. Activation of CD4 T cells promotes the recruitment of LMM A3G into 5- to 15-MDa HMM complexes whose function is unknown. Using tandem affinity purification techniques coupled with MS, we identified Staufen-containing RNA-transporting granules and Ro ribonucleoprotein complexes as specific components of HMM A3G complexes. Analysis of RNAs in these complexes revealed Alu and small Y RNAs, two of the most prominent nonautonomous mobile genetic elements in human cells. These retroelement RNAs are recruited into Staufen-containing RNA-transporting granules in the presence of A3G. Retrotransposition of Alu and hY RNAs depends on the reverse transcriptase machinery provided by long interspersed nucleotide elements 1 (L1). We now show that A3G greatly inhibits L1-dependent retrotransposition of marked Alu retroelements not by inhibiting L1 function but by sequestering Alu RNAs in cytoplasmic HMM A3G complexes away from the nuclear L1 enzymatic machinery. These findings identify nonautonomous Alu and hY retroelements as natural cellular targets of A3G and highlight how different forms of A3G uniquely protect cells from the threats posed by exogenous retroviruses (LMM A3G) and endogenous retroelements (HMM A3G).
Conflict of interest statement
The authors declare no conflict of interest.
Figures

References
-
- Sheehy AM, Gaddis NC, Choi JD, Malim MH. Nature. 2002;418:646–650. - PubMed
-
- Mangeat B, Turelli P, Caron G, Friedli M, Perrin L, Trono D. Nature. 2003;424:99–103. - PubMed
-
- Lecossier D, Bouchonnet F, Clavel F, Hance AJ. Science. 2003;300:1112. - PubMed
-
- Harris RS, Bishop KN, Sheehy AM, Craig HM, Petersen-Mahrt SK, Watt IN, Neuberger MS, Malim MH. Cell. 2003;113:803–809. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
